John-Cunningham virus (JCV) has been recognized as a major contributor to progressive multifocal leukoencephalopathy (PML) in humans, especially in immunodeficient patients. While the prevalence rate of PML in human populations could be up to 80%, the occurrence of tumorigenesis is around 0.07%. Thus, we investigated whether crude-oil extracts could synergistically increase the carcinogenesis along with T antigen from JC virus in vitro. Here we analysed the cell cycle distribution and cytotoxicity of T-antigen of JCV transformed and non-transformed cells by flow cytometry and MTS assay. Our data showed that the high concentration of crude-oil extracts could lead to the disruption of the cell cycle in both T antigen-transformed and non-transformed cells, leading to a G0/G1 arrest. Moreover, the crude-oil extracts could dose-dependently increase cell death and the release of free radicals, which might subsequently result in DNA damage, shown as the expressions of γH2AX proteins. Overall, our data preliminarily revealed that crude-oil extracts could cause the abnormality of non-transformed cells and synergistically induce the possibility of carcinogenesis along with the T antigen of JC virus.
JC virus, Polycyclic aromatic hydrocarbons, Carcinogenesis, DNA damage
John Cunningham virus, as known as JC virus (JCV), is a member of the Polyomaviridae family which is prevalent in the human population. Epidemic research indicates that more than 70% of adults are productively infected by JCV [1-3]. In 1971, JCV was identified as a major viral cause of progressive multifocal leukoencephalopathy (PML) in humans [4]. PML is a fatal progressive inflammation of the white matter of human brains, causing multiple lesions which subsequently lead to the neurological symptoms such as changes in vision, speech, and sometimes personality [5]. However, more studies have suggested that JCV may be associated with tumorigenesis as the T antigens of JCV can transform the host cells, prolong the life span of cells, increase cell proliferation, and facilitate the formation of dense foci in culture [6,7]. Animal studies have also indicated that mice inoculated with JCV would develop multiple tumors in areas including bone, kidney, and subcutaneous connective tissues [8,9]. Despite the high infection rate of JCV in human populations the occurrence rate of tumors is relatively rare. As a result, we contemplate if environmental factors may be important in triggering and exacerbating the process of tumorigenesis. The incidence of oil spills often contaminates the soil and water and causes devastating consequences, including jeopardize the health of human population [10]. A study has clearly indicated that oil spillage is associated with the increased risk of tumorigenesis [11]. Therefore, we aimed to investigate the correlation between crude oil extracts and JCV [12]. In this study, we transfected the fibroblast cells with T antigens obtained from JCV and co-cultured with crude oil extracts to investigate the role of oil spillage attribution to carcinogenesis.
First, we examined whether the T antigen of JCV was transfected into the fibroblast cells [13]. As shown in Figure 1, the photos taken by fluorescent microscopy revealed the expression of T antigen in the nuclei of fibroblast cells, and the transfection of T antigen was also confirmed by Western blotting analysis, indicating the fibroblast cells were transfected with T antigen of JCV. Utilizing these T antigen-transfected cells, we proceeded to test the effects of crude-oil extracts isolated with DMSO from crude oil. Cells were directly co-cultured with different concentrations of from crude-oil extracts for 24, 48, and 72 hours following the MTS assay (Promega®, WI. USA) to quantify the cell viability. Similar to MTT assay, MTS assay is an alternative way to determine the cellular viability based on the activity of mitochondrial dehydrogenase [14]. The data in Figure 2A revealed that the highest concentration (1/50) of crude-oil extract treatment induced significant cytotoxicity over the incubating period similar to the chemotherapy agent, cisplatin, treated cells, whereas the other diluted concentrations of crude-oil extracts (1/100 and 1/500) were less cytotoxic. The rate of viable cells after treatment with 1/500-diluted crude-oil extracts, particularly, remained the same through 72 hours incubation, suggesting that the crude-oil extracts could have mild toxicity when treating fibroblast cells with moderate concentrations. To further understand if the cell death resulting from crude-oil extracts was associated with apoptosis, we explored the cell cycle distribution of T antigen-transfected cells with flow cytometry. The results showed that the cells treated with 1/50 and 1/100 dilutions of crude-oil extracts exhibited higher cell populations in G0/G1 phase, indicating these cells underwent G0/G1 arrest and the cell death could be due to apoptosis after crude-oil extract treatment (Figure 2B).
 Figure 1: The transfection of T antigen of JCV to fibroblasts. The cells were transfected with the plasmid containing T antigen gene from JCV. The T antigen expression was evaluated by staining with anti-T antigen antibody and visualized with fluorescent microscopy. Secondary antibody conjugated with AF488 was applied following the anti-T antigen antibody. DAPI was used for nucleus staining. Photos were taken at magnification 400X.
View Figure 1
Figure 1: The transfection of T antigen of JCV to fibroblasts. The cells were transfected with the plasmid containing T antigen gene from JCV. The T antigen expression was evaluated by staining with anti-T antigen antibody and visualized with fluorescent microscopy. Secondary antibody conjugated with AF488 was applied following the anti-T antigen antibody. DAPI was used for nucleus staining. Photos were taken at magnification 400X.
View Figure 1
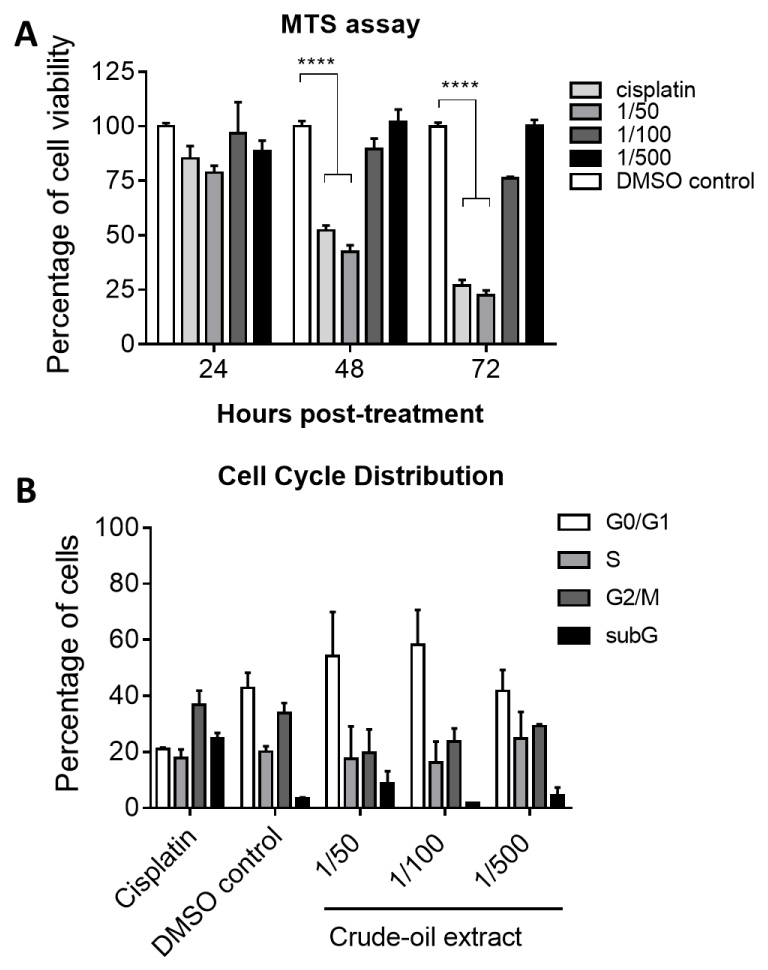 Figure 2: Cell viability and cell cycle distribution after crude-oil extracts treatments. The T antigen-transfected cells were co-cultured with the indicated concentrations of crude-oil extracts for up to 72 hours prior to performing MTS assay and stained with propidium iodide for analysing DNA contents. The cell cycle distribution analysis was conducted with the Guava® easyCyte flow cytometer (Merck-Millipore, MA. USA). ****indicates p-value is less than 0.0001.
View Figure 2
Figure 2: Cell viability and cell cycle distribution after crude-oil extracts treatments. The T antigen-transfected cells were co-cultured with the indicated concentrations of crude-oil extracts for up to 72 hours prior to performing MTS assay and stained with propidium iodide for analysing DNA contents. The cell cycle distribution analysis was conducted with the Guava® easyCyte flow cytometer (Merck-Millipore, MA. USA). ****indicates p-value is less than 0.0001.
View Figure 2
DNA damage usually came along with apoptosis [15,16], and the crude-oil extracts have been proven to generate free radicals, such as reactive oxygen species (ROS), and subsequently induce DNA damage [17,18]. To further investigate whether the crude-oil extracts could result in DNA damages, we stained the DNA damage marker, γH2AX protein, with anti-γH2AX antibody and fluorescent microscopy to visualize the DNA damages on crude-oil-treated cells. Compared with the positive control of cisplatin-treated cells which has been shown to trigger DNA damage in cancer cells [19,20], the fluorescent signals of γH2AX protein in the 1/50 crude-oil extract treatment group were more remarkable than the DMSO control (Figure 3A). Also, we measured the concentrations of ROS with or without crude-oil extract treatment by utilizing the fluorescent dye, H2DCFDA (Thermo Fisher Scientific, PA. USA) [21,22]. To our surprise, only the 1/50 crude-oil extract treatment generated the comparable ROS similar to cisplatin but not the other diluted from crude-oil extract groups (Figure 3B), suggesting that the ROS might play a partial role in mediating the cell death or DNA damage caused by crude-oil extracts. Lastly, to consolidate the potential of carcinogenesis of crude-oil extracts in T antigen-transfected cells, we proceeded to perform the clonogenic assay, an in vitro approach to evaluate the ability of possible carcinogenic agents to induce unlimited division in treated cells. We cultured both T antigen-transfected and untransfected fibroblast cells for 6 passages continuously with or without crude-oil extracts in the culture medium prior to monitoring the colony numbers. As the results in Figure 4 show, the crude-oil-extract treated fibroblast cells regardless of T antigen exhibited more colony numbers and bigger sizes (Figure 4A). Cells transfected with T antigen from JCV promoted the ability of colony formation of fibroblast cells by 51.4% on average in the absence of crude-oil extracts, and the crude-oil extract treatment on the untransfected cells increased the colony formation by 118.1% compared to non-crude-oil extract treated cells. Meanwhile, the colony numbers were raised to 147.2% in T antigen-transfected cells along with long-term crude-oil extract treatment. As summarized in Figure 4B, the data indicate that the combinational stimuli from T antigen and crude-oil extracts could play out a synergistic effect on boosting the colony formation, suggesting the potential of crude-oil extracts to induce carcinogenesis in T antigen-transfected cells.
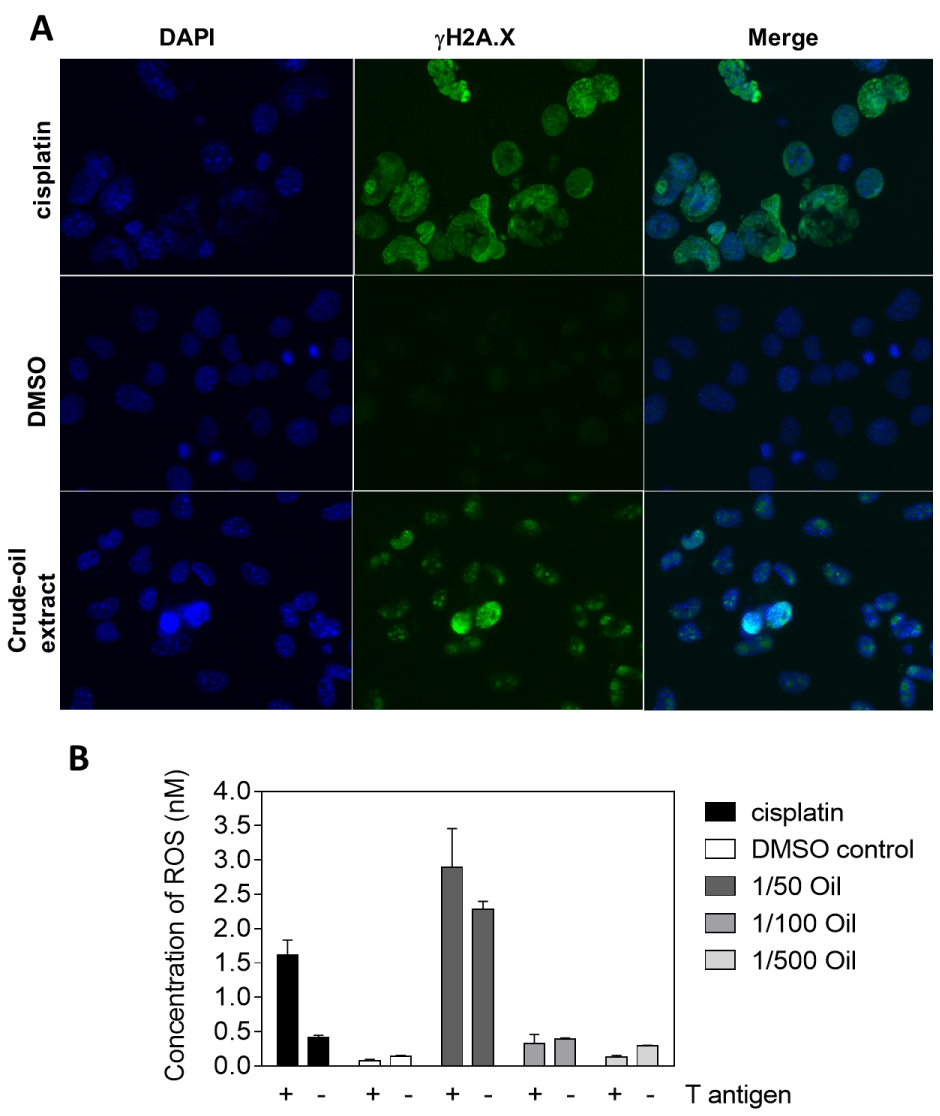 Figure 3: DNA damage and reactive oxygen species induced by crude-oil extracts treatments. A) The DNA damage marker, γH2AX, of T antigen transfected cells was visualized with fluorescent microscopy after 72 hours of crude-oil extracts incubation. Cisplatin treatment was applied as a positive control to justify the γH2AX expression. Magnification was 400X; B) The ROS generation was determined by ELISA reader after adding the fluorescent dye, H2DCFDA, to crude-oil-treated T antigen transfected- cells for 72 hours.
View Figure 3
Figure 3: DNA damage and reactive oxygen species induced by crude-oil extracts treatments. A) The DNA damage marker, γH2AX, of T antigen transfected cells was visualized with fluorescent microscopy after 72 hours of crude-oil extracts incubation. Cisplatin treatment was applied as a positive control to justify the γH2AX expression. Magnification was 400X; B) The ROS generation was determined by ELISA reader after adding the fluorescent dye, H2DCFDA, to crude-oil-treated T antigen transfected- cells for 72 hours.
View Figure 3
 Figure 4: Synergistic effects of crude-oil extracts and T antigen on the ability of colony formation. T antigen-transfected and untransfected cells were treated with or without crude-oil extracts in the culture medium for 6 passages before counting the colonies. A) The representative photos of colony formation with or without T antigen and crude-oil extracts; B) The quantified data. Two-way ANOVA was performed to evaluate the statistical significance.
Figure 4: Synergistic effects of crude-oil extracts and T antigen on the ability of colony formation. T antigen-transfected and untransfected cells were treated with or without crude-oil extracts in the culture medium for 6 passages before counting the colonies. A) The representative photos of colony formation with or without T antigen and crude-oil extracts; B) The quantified data. Two-way ANOVA was performed to evaluate the statistical significance.
*indicates the differences between groups are statistically significant.
View Figure 4
In summary, our findings in this study reveal that crude-oil extracts from contaminated environment could increase the cell turnover rate, facilitate DNA damage, and promote moderate free radical production along with the T antigen from JCV transfected cells, all of which potentially contribute to the increased ability of colony formation, possibly the starting point of carcinogenesis. Overall, our study will not only inspire more attention to focus on the impacts of oil spills or leaks of vapor-phase gasoline on our health and community but also emphasize the potential concerns of neglected JCV in public health.