Vildagliptin is one of the Dipeptidyl Peptidase-4 (DPP-4) inhibitors. It is used as a novel oral anti-diabetic drug for the treatment of Type 2 Diabetes Mellitus (T2DM) with significant control of glycated hemoglobin and a lower incidence of hypoglycemia or weight gain.
The present study hypothesized that the DPP-4 inhibitor vildagliptin may exert protective effects on liver in Non-Alcoholic Fatty Liver Disease (NAFLD) in cholesterol-fed albino mice.
Thirty six albino mice were divided into 3 group (n=12). Group 1 (control) received distilled water and fed normal diet. Group 2 fed high cholesterol diet without therapy. The third group was fed with cholesterol rich diet and daily treated with vildagliptin (15 mg/kg/day for 12 weeks). Liver index, liver triglyceride contents, serum hepatic transaminases, some anti-oxidant enzymes and TNF-α levels were measured in all groups.
Administration of vildagliptin in cholesterol-fed mice attenuated the development of NAFLD via a significant (p < 0.05) decrease in ALT & AST liver enzymes versus NAFLD non-treated group (33.43 ± 6.0 & 34.13 ± 4.7U/L versus 344.9 ± 12.1 & 278.9 ± 5.9 respectively). Additionally, both liver triglyceride contents and tumor necrosis factor-alpha (TNF-α) were significantly (p < 0.05) reduced in treated group 3 compared to cholesterol-fed non-treated groups (for liver triglycerides: 12.61 ± 0.41 versus 56.41 ± 2.11 mg/g tissue protein respectively while for TNF-α: 1.25 ± 0.75 versus 10.78 ± 1.18 pg/mL respectively). Meanwhile, it produced a significant (p < 0.05) increase in the levels of measured serum antioxidant markers in group 3 versus group 2 as follow: (SOD enzyme: 37.5 ± 4.24 versus 0.84 ± 0.43IU/mL, TBARS: 3.46 ± 0.66 versus 62.5 ± 3.3nmol/mg hepatic tissue protein, Catalase enzyme 54.7 ± 3.45 versus 4.51 ± 0.32 and GPx enzyme 17.34 ± 0.79 versus 0.21 ± 0.02 unit/mg hepatic tissue protein, respectively).
These results suggest possible anti-oxidant and partial anti-inflammatory mechanisms in therapy of NAFLD by this anti-diabetic drug.
Vildagliptin, Non-alcoholic fatty liver, Liver index, Liver triglyceride contents, TNF-α, Antioxidant markers
Non-alcoholic fatty liver disease is a leading cause of end stage liver cell failure. This condition is characterized by a progression from simple steatosis to Non-Alcoholic Steatohepatitis (NASH) then to fibrosis [1].
Experimental studies report that in cases of insulin resistance, there is an accumulation of fat in hepatocytes that induces the process of oxidative stress in hepatic cells. Oxidative markers, produced during this kind of stress, activate an inflammatory cascade that results in hepatocellular necrosis [2,3].
There is a close association between inflammation and development of the pathological process of NAFLD [4,5]. Additionally, there is an up-regulation of gene expression of TNF-α with the appearance of repeated cycles of oxidative stress and their markers [6,7].
Although there is many trials to use antioxidant drugs in treatment of NAFLD but there is no clear and beneficial therapeutic strategy till now for this critical condition.
Molecular studies proved that there is an impairment of insulin signaling and a reduction of insulin sensitivity as molecular causes for steatosis. Insulin resistance is associated with a defect in insulin signaling mechanism and results in metabolic disorders in both glucose and lipid metabolism. Insulin resistance appears before type 2 diabetes mellitus and the strong association between hepatic steatosis and insulin resistance has been properly reported [8,9].
Oxidative stress associated with mitochondrial dysfunction in cases of steatosis is strongly related to an increase in serum TNF-α, insulin resistance, body mass index (BMI) values and a progression in fatty acid oxidation [10].
Vildagliptin, one of the Dipeptidyl Peptidase-4 (DPP-4) inhibitors, is used as a novel oral anti-diabetic drug for the treatment of Type 2 Diabetes Mellitus (T2DM). It possesses a potential beneficial role in nephro protection via an anti-oxidant and anti-apoptotic effects in type 2 diabetic patients [11,12].
The present study aims to investigate the possible protective effect on liver homogenates that would be induced by vildagliptin in a model of non-alcoholic fatty liver disease in cholesterol-fed albino mice for 12 weeks.
Vildagliptin powder was obtained from Shengda Pharmaceutical Co., Limited and dissolved in distilled water (DW) (Hong-kong, Japan). Cholesterol powder was purchased from Sigma chemical company, prepared as 2% (w/w) and dissolved in DW (St Louis, MO, USA). Serum alanine & aspartate transaminases (ALT, AST) spectrum diagnostic kits and triglycerides kit were obtained from Biodiagnostics, Cairo, Egypt. All other chemicals and kits are purchased from Sigma chemical company (St Louis, MO, USA).
Albino mice weighing 20-25 g were placed in labomiceory of Pharmacology dept., Faculty of medicine, Ain Shams University. They were housed in cages with a natural light-dark cycle and fed on a standard pellet diet and water ad libitum. All procedures were in accordance with the National Institute of Health's Guide for the Care and Use of Labomiceory Animals, as well as the guidelines of the Animal Welfare Act.
Thirty-six albino mice were left in the labomiceory for one week for acclimatization. Then, they were assigned into three experimental groups (n=12 mice/group)
Group 1: control group with normal fed diet received distilled water per os (po) daily for 12 weeks. Induction of hypercholesterolemia in groups 2 and 3 according to Csont [13]
Group 2: mice on labomiceory chow enriched with 2% (w/w) cholesterol, received distilled water po daily for 12 weeks as positive control group
Group 3: mice, on labomiceory chow enriched with 2% (w/w) cholesterol, received vildagliptin dissolved in distilled water in a dose of 15 mg/kg po daily for 12 weeks. Dose was used according to a pilot study done by the author before this present study.
Mice were fasted for 12 hours before anesthesia and blood collection. At the end of the 12th week, all tested mice were anesthetized by urethane (1g/kg) intraperitoneally (ip). They were dissected and blood samples were collected from abdominal aorta and prepared for biochemical analysis. Serum was obtained from each sample by centrifugation of the clotted blood. Then, their livers were rapidly removed and homogenized for further biochemical tests. Dead bodies of mice were burn in its specific site in the animal house of Faculty of Medicine, Ain Shams University.
NB: A pilot study, done on a group of mice (n=12) that was injected with 1g/Kg urethane ip, showed no significant differences in the measured parameters of the present study from the control group 1.
Triglycerides were assayed in hepatic tissue using commercially available kit after lipid extraction [14].
Serum levels of Alanine and Aspartate Transaminases (ALT & AST): They were measured using biochemistry automatic analyzer (Hitachi 7600)
Determination of SOD enzyme level in erythrocyte lysates: At the end of the 12th week of the study, blood samples were collected from abdominal aorta of dissected tested mice of all groups for measurement of SOD levels in erythrocyte lysates, using commercially-available colorimetric assay kits, based on an indirect xanthine-xanthine oxidase method as described by Sato et al. [15] and results were expressed in IU/mL.
Hepatic lipid peroxidation assay as Thiobarbituric Acid-Reactive Substance (TBARS): Hepatic lipid peroxidation was quantified by measuring thiobarbituric acid-reactive substance (TBARS) according to Fraga et al. [16]. Liver tissue of each mouse was homogenized in 9 volumes of 50mmol/L Tris-HCl buffer (pH 7.4) containing 180mmol/L KC1, 10mmol/L EDTA and 0.02% butylated hydroxytoluene. 0.2ml of 8.1% sodium dodecyl sulfate, 1.5ml of 20% acetic acid, 1.5mL of 0.9% thiobarbituric acid, and 0.6 ml of distilled water were added to 0.2 ml of the tissue homogenate and vortexed. The reaction mixture was placed in a water bath at 95℃ for 1 hour. After cooling on ice, 1.0ml of distilled water and 5.0mL of butanol/pyridine mixture (15:1, v/v) is added and vortexed. After centrifugation at 10,000 x g for 10 minutes, absorbance of the resulting precipitate was determined at 532 nm. The TBARS concentration was calculated using 1,1,5,5-tetraethoxypropane as standard.
Determination of catalane enzyme activity: Catalase (CAT) activity in the liver homogenates was assayed colorimetrically as described by Sinha [17] using dichromate-acetic acid reagent 5% potassium dichromate and glacial acetic acid were mixed in 1:3 ratio. The intensity was measured at 620 nm and the amount of hydrogen peroxide hydrolyzed was calculated for the catalase activity .
Determination of Glutathione Peroxidase [GPx] enzyme activity: Glutathione Peroxidase (GPx) activity in the liver homogenates was measured by the method described by Rotruck et al. [18]. Glutathione Peroxidase (GPx) was assayed in the liver sample homogenized in 8 volumes of cold buffer (50mM Tris-HCl, pH 7.5, containing 5mM EDTA and 1 mM 2-mercaptoethanol), next centrifuged 8500xg for 10 minutes at 40℃ GPx activity was determined in supernatant using BIOXYTECH GPx-340TM Assay kit produced by OXIS International, Inc., USA. The GPx assay was based on the oxidation of NADPH to NADP+, which is accompanied by a decrease in absorbance at 340nm. The mice of this decrease are directly proportional to the GPx activity in the sample.
Measurement of hepatic tumor necrosis factor-alpha (TNF-α): TNF-α was measured by a quantitative sandwish enzyme immunoassay technique with microplates pre-coated with a mice TNF-α specific monoclonal antibody (Mice TNF-αlpha ELISA kit). We followed the instructions written in the commercial kit [19].
Protein determination: The protein content of liver homogenates was determined by spectrophotometer according to the method of Bradford [20]. The aim is to relate TBARS concentration as nmol/mg tissue protein, CAT and GPx enzyme activities as unit/mg tissue protein.
Results are expressed as mean ± SD [Standard Deviation]. Statistical analysis was performed by analysis of variance followed by Tukey's post hoc using GraphPad Prism version 3.00 for Windows 97 (Graph Pad Software, San Diego, CA, U.S.A.). Differences with p< 0.05 were considered to be statistically significant.
Table 1 showed significant (p< 0.05) increase in both serum AST & ALT in cholesterol-fed mice group compared with control group. Both enzymes were decreased to normal values in vildagliptin-treated group.
Table 1: Significant (p< 0.05) increase in both serum AST & ALT in cholesterol-fed mice group compared with control group. View Table 1
p< 0.05= significant increase in liver enzymes in cholesterol-fed non-treated group compared to the control non-treated albino mice group.
p< 0.05= significant reduction in liver enzymes in vildagliptin-treated group compared to the cholesterol fed -non treated albino mice group
Table 2 shows Liver contents of triglycerides were significantly increased (p< 0.05) in cholesterol-fed non-treated group compared with control group. Vildagliptin-treated group showed a significant (p< 0.05) reduction in liver triglycerides compared to the other tested groups.
Table 2: Liver contents of triglycerides were significantly increased (p< 0.05) in cholesterol-fed non-treated group compared with control group. View Table 2
p< 0.05= significant increase in liver triglycerides in cholesterol-fed non-treated group compared to the control non-treated albino mice group
p< 0.05 = significant reduction in liver triglycerides in vildagliptin-treated group compared to cholesterol fed - non treated albino mice group
Effect of 12-weeks administration of tested drug on levels of Superoxide Dismutase [SOD] enzyme in IU/mL in erythrocyte lysates of the tested albino mice. Results are expressed as mean ± SD (n=12 mice/group) (Figure 1)
 Figure 1: Effect of vildagliptin on levels of Superoxide Dismutase enzyme [SOD] in erythrocyte lysates of different groups in IU/mL. View Figure 1
Figure 1: Effect of vildagliptin on levels of Superoxide Dismutase enzyme [SOD] in erythrocyte lysates of different groups in IU/mL. View Figure 1
Significant (*P< 0.05) increase in SOD levels was obtained with Group 3 (treated with vildagliptin) when compared to Group 2 (cholesterol-fed non treated mice).
p< 0.05= significant decrease in SOD enzyme levels in group 2 compared to the control non-treated albino mice group 1 p< 0.05 = significant increase in SOD enzyme levels in vildagliptin-treated group 3 compared to the cholesterol fed -non treated albino mice group 2
Results are expressed as mean ± SD (n=12 albino mice/group).
A significant (P < 0.05) decrease in TBARS levels in comparison to the cholesterol-fed non treated Group 2. The TBARS levels in treated group (3) were comparable to that measured in the control group 1.
p< 0.05= significant increase in TBARS levels in group 2 compared to the control non-treated albino mice group 1(Figure 2) p< 0.05 = significant decrease in TBARS levels in vildagliptin-treated group 3 compared to the cholesterol- fed non treated albino mice group 2
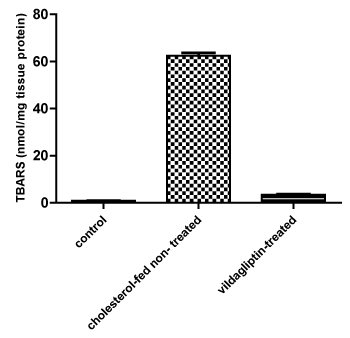 Figure 2: Effect of 12-weeks administration of vildagliptin on Thiobarbituric Acid-Reactive Substance (TBARS) in nmol/mg tissue protein of the liver tissue homogenates of the tested Mice. View Figure 2
Figure 2: Effect of 12-weeks administration of vildagliptin on Thiobarbituric Acid-Reactive Substance (TBARS) in nmol/mg tissue protein of the liver tissue homogenates of the tested Mice. View Figure 2
A significant decrease in the activities of these enzymes was noted in cholesterol-fed non-treated mice. Vildagliptin-treated group showed that their activities were significantly increased compared to cholesterol- fed non-treated group (Table 3).
Table 3: Mean ± SD changes in the activities of liver CAT, GPx in control and experimental albino mice expressed as unit/mg tissue protein of liver homogenates. View Table 3
There is a significant (p< 0.05) increase in hepatic TNF-α in cholesterol-fed non-treated group compared to control group. While a significant (p< 0.05) decrease in its level in hepatic tissue of albino mice treated with vildagliptin for 12 weeks.
Figure 3 shows effect of 12-weeks administration of vildagliptin on hepatic TNF-α (pg/mL) of the tested albino mice. Results are expressed as mean ± SD (n=12 albino mice/group).
 Figure 3: Effect of 12-weeks administration of vildagliptin on hepatic TNF-α (pg/mL) of the tested albino mice. View Figure 3
Figure 3: Effect of 12-weeks administration of vildagliptin on hepatic TNF-α (pg/mL) of the tested albino mice. View Figure 3
A significant (*P< 0.05) increase in hepatic TNF-α levels in comparison to the model untreated Group 2. The hepatic TNF-α level in treated group 3 was comparable to that measured in the control group 1.
p< 0.05= significant increase in hepatic TNF-α levels in group (2) compared to the control non-treated albino mice group 1 p< 0.05 = significant decrease in hepatic TNF-α levels in vildagliptin-treated group 3 compared to the cholesterol- fed non treated albino mice group 2
Vildagliptin, one of the Dipeptidyl Peptidase-4 (DPP-4) inhibitors and used as a novel oral anti-diabetic drug for the treatment of Type 2 Diabetes Mellitus (T2DM), reversed the changes induced by experimental induction of steatohepatits in the laboratory markers, the targets of the present study, to levels that are almost comparable to the control group.
The present study demonstrated that steatohepatitis was induced by 2% cholesterol diet for albino mice as evidenced by a significant increase in hepatic tissue content of triglycerides and serum levels of ALT &AST hepatic enzymes.
The increase in the hepatic tissue content of triglycerides accompanied by hepatic lipid peroxidation and an increase in the availability of inflammatory markers are the main causes of hepatic injury in cases of NAFLD [21].
Similar findings were reported by Xu et al. [22] who worked on Pioglitazone (PGZ) as a Peroxisome Prolifemiceor Activated Receptor Gamma (PPARγ) used as an oral anti-diabetic drug. It was found in this study that PGZ treatment group decreased the elevation of serum levels of ALT, ALP, TNF-α and hepatic injury-induced any Ischemia-Reperfusion (IR) technique in high fat-fed mice with NAFLD.
Additionally, leakage of serum transaminases (ALT & AST) lead to massive injury of hepatic cells that plays a major role in NAFLD [23]. A single oral daily dose administration of vildagliptin, in the present study, approximately normalized the levels of these enzymes to be comparable to the control group.
On the other hand, the increase in hepatic content of triglycerides, reported in the present study, could be explained by failure of the liver of cholesterol-fed mice to synthesize apolipoprotein that is essential for quenching and transporting triglycerides from the liver according to Thong-Ngam et al. [24].
As regard its mechanism against oxidative stress, vildagliptin reversed the increase in TBARS as a marker of hepatic lipid peroxidation in the present study. This could point to an anti-oxidant effect of the drug especially with the increase in SOD anti-oxidant enzyme in erythrocyte lysates together with catalase & glutathione peroxidase enzyme activities of tested cholesterol-fed albino mice. This beneficial anti-oxidative effect of vildagliptin against lipid peroxidation is considered to be a major protective factor against oxidative damage in serious diseases [25].
A study reported that the reduction in hepatic lipid peroxidation by diet regulation exerts a high beneficial response on hepatic and the whole body anti-oxidative response especially in cases of NAFLD [26].
Another study demonstrated that plasma non-esterified fatty acid are exposed to a high rate of their uptake into hepatic cells in parallel to an increase in their release from adipose tissue due to a reduction in activity of insulin in cases of NAFLD. These cases suffer from hyperinsulinemia that increases the up regulation of genes together with a promotion of de novo lipogenesis in the liver. Insulin resistance in NAFLD was found to be related to oxidative stress and it exerts a significant increase in hepatic lipid accumulation with disturbance in hepatic triglyceride homeostasis contributing to the pathogenesis of NAFLD. However, the significant reduction of hepatic accumulation of triglycerides with the significant increase in activities of hepatic antioxidant enzymes by vildagliptin in NAFLD model treated mice could be related to its ability to counteract hyperinsulinemia and de novo lipogenesis in hepatic cells by oxidative stress. These actions would provide a hepatoprotective ability of vildagliptin in NAFLD [27]. This was similar to reported results obtained from the administration of thioglitazone e.g. pioglitazone or telmisartan as an Angiotensin Receptor Blocker (ARB). These results pointed to the abilities of such drugs to reverse insulin resistance with a reduction in circulating bad lipids leading to protection against accumulation of triglycerides in hepatic tissue. These beneficial effects were suggested to be due to the action of pioglitazone on PPAR-PPARγ receptors which are involved in the metabolism of carbohydrates, lipids and inflammatory process and telmisartan on PPAR-a receptors which are also involved in its dyslipidemic action with reduction in synthesis of esterified fatty acids and prevention of accumulation of hepatic triglycerides [28].
Additionally, the increase in hepatic content of TNF-α and other proinflammatory cytokines could be related to the inflammatory process related to the development of steatohepatitis up to NAFLD as reported by Paschos and Paletas [29] & Kamal [30].
This research was officially supported by the Medical Research Service of the Ain Shams University. It was financially supported by the laboratory of the Pharmacology Department, Faculty of Medicine, Ain Shams University.