

International Journal of Neurology and Neurotherapy
Cold Shock as a Possible Remedy for Neurodegenerative Disease
Takuma Aihara1 and Fuminori Tsuruta1,2*
1Graduate School of Life and Environmental Sciences, University of Tsukuba, Japan
2Ph.D. Program in Human Biology, School of Integrative and Global Majors, University of Tsukuba, Japan
*Corresponding author:
Fuminori Tsuruta, Graduate School of Life and Environmental Sciences, University of Tsukuba, 1-1-1 Tennodai, Tsukuba, Ibaraki 305-8577, Japan, Tel: +81-29-853-6887, E-mail: tsuruta.fuminori.fn@u.tsukuba.ac.jp
Int J Neurol Neurother, IJNN-3-053, (Volume 3, Issue 4), Mini Review; ISSN: 2378-3001
Received: May 18, 2016 | Accepted: July 20, 2016 | Published: July 22, 2016
Citation: Aihara T, Tsuruta F (2016) Cold Shock as a Possible Remedy for Neurodegenerative Disease. Int J Neurol Neurother 3:053. 10.23937/2378-3001/3/4/1053
Copyright: © 2016 Aihara T, et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Abstract
Synapse remodeling is an essential physiological phenomenon in maintaining the normal brain function. Traumatic injury is one of the causes that triggers abnormality in synapse remodeling, leading to neurodegeneration. So far, substantial clinical trials have been attempted to cure neurodegeneration. One of the challenging approaches that improves neurodegenerative symptoms is therapeutic hypothermia. The putative positive feature induced by therapeutic hypothermia on patient is neuroprotection; suppressing neuronal apoptosis and inflammatory response. Recently, it has been reported that RBM3 (RNA binding motif protein 3), which is a cold shock protein expressed in brain when mammals were exposed to the cold temperature, negatively regulates the progression of neurodegeneration. However, the precise mechanism how this protein acts on brain is not fully understood. In this review, we summarize the possible mechanism of downstream pathway induced by therapeutic hypothermia associated with neuroprotection.
Keywords
Synapse remodeling, Therapeutic hypothermia, RBM3, Low metabolism, Autophagy
Introduction
The normal brain function is tightly maintained by the elimination and reformation of synapse, which creates meticulously organized synapse network [1]. Incessant synapse loss by traumatic injury or by aging could result in neurodegenerative diseases, such as Alzheimer disease and Parkinson disease [2,3]. There are numerous publications revealing the mechanism of synapse loss, although less is known about how synapse is reformed. There have been several attempts to establish an efficient treatment for neurodegenerative diseases, but still the number of treatment is insufficient.
Although efficient treatment for neurodegenerative disease is limited, downstream pathway induced by therapeutic hypothermia is thought to play a positive effect on suppressing the ongoing exacerbation. One of the possible key features of therapeutic hypothermia is neuroprotection [4]. For instance, after the traumatic injury in brain, neuronal apoptosis and inflammation pathway are activated, resulting in further exacerbation [5,6]. Inducing therapeutic hypothermia could suppress these ongoing reactions [4]. The mechanism of how therapeutic hypothermia could suppress the ongoing neurodegeneration is inspired by hibernating animals. Dynamic synapse remodeling and morphological changes in neuron are observed in these animals along with the change in body temperature during torpor and waking state [7-9]. When animals were exposed to cold temperature, synaptic contacts get reduced, while rewarming forms new synaptic connections [7-9]. The exposure to cold temperature triggers the expression of RBM3 (RNA binding motif protein 3), which takes part in neural plasticity, suppresses the synapse loss derived from neurodegenerative diseases [9-11]. The precise mechanism how this protein acts on brain has not been elucidated. Hence, further understanding toward downstream pathway induced by therapeutic hypothermia could be a breakthrough in neurodegenerative disorder treatment. In this review, we propose a clue in neurodegenerative disease by targeting cold shock as a possible remedy.
Therapeutic Hypothermia
Immediate treatment after the brain damage is required to prevent further exacerbation. Therapeutic hypothermia is one of the candidates thought to be a potential approach in suppressing neuronal apoptosis, inflammatory response and oxidative stress [4]. Although several attempts have been made to demonstrate the possible effect of therapeutic hypothermia in clinical trial, many studies failed to show a dramatic effect due to the unestablished way of conducting therapeutic hypothermia safely and effectively on patients [12,13]. In addition, how the downstream pathway induced by therapeutic hypothermia is involved in palliating neurodegenerative disease is not widely studied. Therefore, an investigation of therapeutic hypothermia is a challenging trial to explore a novel medical approach. Theoretically, it is said that an operation of therapeutic hypothermia between the range of 34-36°C is the optimum temperature to suppress the exacerbation of brain damage [14,15]. There are several ways to trigger therapeutic hypothermia, and major approaches are whole-body surface cooling and endovascular cooling [14]. In order to prevent shivering, patients are usually anesthetized [16]. When aiming at the certain parts of the brain only, cooling caps and helmets are common way, although further research must be conducted to support the efficacy for this method [14]. It should be stated that inducing excessively low body temperature abruptly on patient increases the tissue damage. Hence, slow induction of optimal body temperature for the clinical trial is necessary.
Patient with induced therapeutic hypothermia shows several physiological changes. The drop in heart rate and blood pressure could be observed. Respiration rate and metabolic rate significantly drop. Interestingly, similar phenomena are observed in hibernating animals. Various mammalian species undergo hibernation in order to endure the harsh environment when nourishments are scarce. During hibernation, their metabolic rate significantly drops to as low as 1% of active state [17]. The heart rate and blood pressure decrease, as well as gas exchange rate drops [18,19]. The blood flow to the central nervous system (CNS) gets suspended [14,20]. In general, lack of blood flow in CNS results in a critical condition for human and other mammals. The interruption of the blood flow, as well as nutrient and oxygen supply, provokes malfunction in brain. Interestingly, the synapse loss could be observed by the drop of body temperature, but reformation takes place by rewarming [7-9]. It is still not clear how hibernating animals can withstand the significant drop of body temperature and metabolism. In addition, how animals survive by the drastic reduction of blood flow in CNS, without damaging the brain function, is yet to be elucidated. Thus, how synapse remodeling is controlled by the temperature change was an enigma, but recent report showed that RBM3 plays a key role in this regulation. RBM3 is said to be expressed at dendrites in hippocampus, suppressing neural apoptosis and synapse elimination [11,21,22]. In practice, conducting early cooling on Alzheimer mice and prion-infected mice prolong the surviving date, detecting more RBM3 [9]. Further study about RBM3 can bring new therapeutic targets for neurodegenerative disease. Moreover, elucidating the mechanism of hibernation and inducing hibernation-like state on patients could be a treatment for several neurodegenerative disorders, resulting in a low mortality rate.
Induction of Low Metabolism
In nature, environmental light and temperature have a close linkage. An exposure to a constant darkness is regarded as one of the factors in triggering torpor by modulating the circadian rhythm [23,24]. It has been reported that animals under the condition of 12 h dark/12 h dark has a lower body temperature and metabolism compared to the animals placed under the condition of 12 h light/12 h dark [25]. The pathways that regulate body temperature and energy metabolism are both located in hypothalamus with the connection of nuclei. Animal experienced constant darkness increases 5'-AMP in blood, a substance that plays a key role as an internal signal indicating that the cells are under metabolic stress [25,26]. Usually, AMP is generated as a by-product from the reaction generating ATP from ADP [27]. Animals use ATP as a primary energy source for sustaining the biofunction. When ATP generation fails to keep pace with ATP utilization, the intracellular concentration of ADP increases in order to generate more ATP. Hence, since animals during hibernation do not generate much ATP compared to their waking state, the ratio of AMP/ATP rises. 5'-AMP directly activates AMP-activated protein kinase (AMPK), which is a molecular stress response pathway [28,29]. The activation of AMPK up-regulates and down-regulates several pathways. For instance, fatty acid oxidation and glucose transport are facilitated, leading to ATP generation [30]. AMPK down-regulates the activation of mammalian target of rapamycin complex 1 (mTORC1), which is a sensor of cellular nutrient status [26,31]. An active mTORC1 during waking state promotes cell growth and proliferation, while it decreases energy-recycling processes such as autophagy [32]. As stipulated previously, blood flow in CNS decreases during hibernation [14,20], resulting in the lack of nutrients and oxygen supply in brain. In order to allow neurons to survive, mTORC1 is suppressed and stimulates autophagy, provides nutrients to the cells [26,31,32]. It is reported that injecting 5'-AMP in laboratory rodents can induce artificial torpor, leading to a drop in body temperature and metabolism [25]. Exposing laboratory rodents to a cold temperature for a certain period without injecting 5'-AMP could cause a severe tissue or organ damage due to the significant drop of the body temperature. However, cooling the animals after the injection of 5'-AMP has little risk in damaging the tissues and organs, having no side effects after the rewarming [33]. Thus, therapeutic hypothermia with 5'-AMP injection to the patient could be a new foray in establishing a safer treatment for neurodegeneration [33].
Autophagy and Neurodegeneration
Cells are constantly undergoing protein production and degradation to maintain homeostasis in brain [34]. Autophagy is a bulk protein degradation pathway triggered by starvation and protein turnover [35,36]. The aggregated abnormal-structured proteins are ubiquitinated, then engulfed by double-membraned vesicles, known as autophagosomes. The fusion of outer membranes of autophagosomes and lysosome results in the degradation of the aggregated proteins by the enzymes derived from lysosome [37-40]. Consequently, newly generated amino acids are reused to maintain the intracellular homeostasis. Recently, it has been reported that aggregation of intracellular misfolded protein due to the abnormality in autophagy features many late-onset neurodegenerations such as Huntington's disease, Alzheimer disease and Parkinson disease [41]. Loss of Atg7, which is an essential enzyme for autophagy, leads to neurodegeneration [42]. Mutant mice gradually show behavioral defects and a reduction in coordinated movement, surviving almost one month of birth [36]. There is a report showing a large reduction in the number of Purkinje cells in ATG7 KO mice, and similar neural loss could be recognized in the hippocampal pyramidal cell layer [36]. It is thought that activating autophagy artificially can cure neurodegeneration derived from abnormal protein aggregation [41]. Since the onset of low body temperature triggers low metabolism and autophagyby the possible activation of AMPK [26,31,32], therapeutic hypothermia could be an effective approach. Moreover, RBM3 expressed by cold shock could possibly contribute in suppressing neuronal cell death derived from autophagy deficiency. In practice, there is a report showing that inducing autophagy by rapamycin, which inhibits mTORC1, in transgenic mice with Huntington disease palliated the symptom [43]. Applying rapamycin to the patient could be a useful tactic, although mTORC1 regulates not only autophagy but also translation of certain proteins and cell division, long-term use of rapamycin is considered perilous, resulting in side effects such as poor wound healing and general immunosuppression [41]. Although it is given that inhibiting mTORC1 activates autophagy, the precise pathway how mTORC1 regulates autophagy remains black box [44,45]. Hence, revealing this unknown pathway and targeting mTORC1 for autophagy induction without the repercussion on other biofunctions could be a breakthrough in neurodegenerative disease treatment.
Conclusion
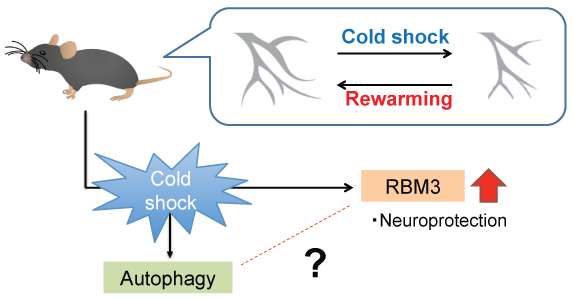
It is certain that downstream pathways induced by therapeutic hypothermia and energy metabolism pathway, as well as autophagy pathway have a close connection. Still the precise mechanism how these pathways are linked together remains unknown. Further study about temperature control, also about RBM3 could give a hint in developing therapeutic hypothermia (Figure 1). Taken together, therapeutic hypothermia on patient is likely to be a promising therapeutic strategy.
References
-
Holtmaat A, Svoboda K (2009) Experience-dependent structural synaptic plasticity in the mammalian brain. Nat Rev Neurosci 10: 647-658.
-
Selkoe DJ (2002) Alzheimer's disease is a synaptic failure. Science 298: 789-791.
-
Lepeta K, Lourenco MV, Schweitzer BC, Martino Adami PV, Banerjee P, et al. (2016) Synaptopathies: synaptic dysfunction in neurological disorders. J Neurochem.
-
Ma H, Sinha B, Pandya RS, Lin N, Popp AJ, et al. (2012) Therapeutic hypothermia as a neuroprotective strategy in neonatal hypoxic-ischemic brain injury and traumatic brain injury. Curr Mol Med 12: 1282-1296.
-
Sahuquillo J, Vilalta A (2007) Cooling the injured brain: how does moderate hypothermia influence the pathophysiology of traumatic brain injury. Curr Pharm Des 13: 2310-2322.
-
Saito K, Fukuda N, Matsumoto T, Iribe Y, Tsunemi A, et al. (2010) Moderate low temperature preserves the stemness of neural stem cells and suppresses apoptosis of the cells via activation of the cold-inducible RNA binding protein. Brain Res 1358: 20-29.
-
Magariños AM, McEwen BS, Saboureau M, Pevet P (2006) Rapid and reversible changes in intrahippocampal connectivity during the course of hibernation in European hamsters. Proc Natl Acad Sci U S A 103: 18775-18780.
-
Popov VI, LS Bocharova (1992) Hibernation-induced structural changes in synaptic contacts between mossy fibres and hippocampal pyramidal neurons. Neuroscience 48: 53-62.
-
Peretti D, Bastide A, Radford H, Verity N, Molloy C, et al. (2015) RBM3 mediates structural plasticity and protective effects of cooling in neurodegeneration. Nature 518: 236-239.
-
Williams DR, Epperson LE, Li W, Hughes MA, Taylor R, et al. (2005) Seasonally hibernating phenotype assessed through transcript screening. Physiol Genomics 24: 13-22.
-
Smart F, Aschrafi A, Atkins A, Owens GC, Pilotte J, et al. (2007) Two isoforms of the cold-inducible mRNA-binding protein RBM3 localize to dendrites and promote translation. J Neurochem 101: 1367-1379.
-
Todd MM, Hindman BJ, Clarke WR, Torner JC, Intraoperative Hypothermia for Aneurysm Surgery Trial (IHAST) Investigators (2005) Mild intraoperative hypothermia during surgery for intracranial aneurysm. N Engl J Med 352: 135-145.
-
Den Hertog HM, van der Worp HB, Tseng MC, Dippel DW (2009) Cooling therapy for acute stroke. Cochrane Database Syst Rev CD001247.
-
Forreider B, Pozivilko D, Kawaji Q, Geng X, Ding Y (2016) Hibernation-like neuroprotection in stroke by attenuating brain metabolic dysfunction. Prog Neurobiol, 30062-3006c9.
-
Schwab S, Schwarz S, Spranger M, Keller E, Bertram M, et al. (1998) Moderate hypothermia in the treatment of patients with severe middle cerebral artery infarction. Stroke 29: 2461-2466.
-
Lyden PD, Hemmen TM, Grotta J, Rapp K, Raman R (2014) Endovascular therapeutic hypothermia for acute ischemic stroke: ICTuS 2/3 protocol. Int J Stroke 9: 117-125.
-
Carey HV, Andrews MT, Martin SL (2003) Mammalian hibernation: cellular and molecular responses to depressed metabolism and low temperature. Physiol Rev 83: 1153-1181.
-
Horwitz BA, Chau SM, Hamilton JS, Song C, Gorgone J, et al. (2013) Temporal relationships of blood pressure, heart rate, baroreflex function, and body temperature change over a hibernation bout in Syrian hamsters. Am J Physiol Regul Integr Comp Physiol 305: R759-768.
-
Milsom WK, Jackson DC (2011) Hibernation and gas exchange. Compr Physiol 1: 397-420.
-
Drew KL, Buck CL, Barnes BM, Christian SL, Rasley BT, et al. (2007) Central nervous system regulation of mammalian hibernation: implications for metabolic suppression and ischemia tolerance. J Neurochem 102: 1713-1726.
-
Chip S, Zelmer A, Ogunshola OO, Felderhoff-Mueser U, Nitsch C, et al. (2011) The RNA-binding protein RBM3 is involved in hypothermia induced neuroprotection. Neurobiol Dis 43: 388-396.
-
Tong G, Endersfelder S, Rosenthal LM, Wollersheim S, Sauer IM, et al. (2013) Effects of moderate and deep hypothermia on RNA-binding proteins RBM3 and CIRP expressions in murine hippocampal brain slices. Brain Res 1504: 74-84.
-
Aschoff J (1960) Exogenous and endogenous components in circadian rhythms. Cold Spring Harb Symp Quant Biol 25: 11-28.
-
Ruby NF, Dark J, Heller HC, Zucker I (1996) Ablation of suprachiasmatic nucleus alters timing of hibernation in ground squirrels. Proc Natl Acad Sci U S A 93: 9864-9868.
-
Zhang J, Kaasik K, Blackburn MR, Lee CC (2006) Constant darkness is a circadian metabolic signal in mammals. Nature 439: 340-343.
-
Young, LH (2008) AMP-activated protein kinase conducts the ischemic stress response orchestra. Circulation 117: 832-840.
-
Hardie DG, Ross FA, Hawley SA (2012) AMPK: a nutrient and energy sensor that maintains energy homeostasis. Nat Rev Mol Cell Biol 13: 251-262.
-
Lindsley JE, Rutter J (2004) Nutrient sensing and metabolic decisions. Comp Biochem Physiol B Biochem Mol Biol 139: 543-559.
-
Weikel KA, Ruderman NB, Cacicedo JM (2016) Unraveling the actions of AMP-activated protein kinase in metabolic diseases: Systemic to molecular insights. Metabolism 65: 634-645.
-
Inoki K, Zhu T, Guan KL (2003) TSC2 mediates cellular energy response to control cell growth and survival. Cell 115: 577-590.
-
Gwinn DM, Shackelford DB, Egan DF, Mihaylova MM, Mery A, et al. (2008) AMPK phosphorylation of raptor mediates a metabolic checkpoint. Mol Cell 30: 214-226.
-
Hosokawa N, Hara T, Kaizuka T, Kishi C, Takamura A, et al. (2009) Nutrient-dependent mTORC1 association with the ULK1-Atg13-FIP200 complex required for autophagy. Mol Biol Cell 20: 1981-1991.
-
Lee CC (2008) Is human hibernation possible? Annu Rev Med 59: 177-186.
-
Forman MS, Trojanowski JQ, Lee VM (2004) Neurodegenerative diseases: a decade of discoveries paves the way for therapeutic breakthroughs. Nat Med 10: 1055-1063.
-
Hara T, Nakamura K, Matsui M, Yamamoto A, Nakahara Y, et al. (2006) Suppression of basal autophagy in neural cells causes neurodegenerative disease in mice. Nature 441: 885-889.
-
Komatsu M, Waguri S, Chiba T, Murata S, Iwata J, et al. (2006) Loss of autophagy in the central nervous system causes neurodegeneration in mice. Nature 441: 880-884.
-
Cuervo AM (2004) Autophagy: in sickness and in health. Trends Cell Biol 14: 70-77.
-
Levine B, Klionsky DJ (2004) Development by self-digestion: molecular mechanisms and biological functions of autophagy. Dev Cell 6: 463-477.
-
Klionsky DJ (2005) The molecular machinery of autophagy: unanswered questions. J Cell Sci 118: 7-18.
-
Mizushima N (2005) The pleiotropic role of autophagy: from protein metabolism to bactericide. Cell Death Differ 12: 1535-1541.
-
Winslow AR, Rubinsztein DC (2008) Autophagy in neurodegeneration and development. Biochim Biophys Acta 1782: 723-729.
-
Ohsumi Y (2001) Molecular dissection of autophagy: two ubiquitin-like systems. Nat Rev Mol Cell Biol 2: 211-216.
-
Ravikumar B, Vacher C, Berger Z, Davies JE, Luo S, et al. (2004) Inhibition of mTOR induces autophagy and reduces toxicity of polyglutamine expansions in fly and mouse models of Huntington disease. Nat Genet 36: 585-595.
-
Davies JE, Sarkar S, Rubinsztein DC (2006) Trehalose reduces aggregate formation and delays pathology in a transgenic mouse model of oculopharyngeal muscular dystrophy. Hum Mol Genet 15: 23-31.
-
Sarkar S, Davies JE, Huang Z, Tunnacliffe A, Rubinsztein DC (2007) Trehalose, a novel mTOR-independent autophagy enhancer, accelerates the clearance of mutant huntingtin and alpha-synuclein. J Biol Chem 282: 5641-5652.
