SARS-COV-2 pandemic is almost 3-years-old and remains a case for concern. An overwhelming production of pro-inflammatory cytokines in the context of COVID-19, not only impairs viral clearance but also promotes paradoxically hyperinflammation including cytokine storm; an unfortunate event leading to organ failure following long term damage due to inflammation and even death.
With the advent of new variants, the interaction between vaccinated and unvaccinated people and also the interaction between persons with different variants or between the healthy and the asymptomatic; all of these possibly leading to the formation of new variants.
CAR T therapy has achieved remarkable clinical results treatment of relapsed/refractory B-cell-derived malignancies.
CAR T and SARS-COV-2 share one thing in common; the cytokine storm even though the former is a treatment associated complication and the latter is an exaggerated host response.
Hence, we propose using Immunopharmacology (which targets amongst others, pathologies in which inflammation is the main component), by using anti-CAR-engineered T to target the cytokine storm in COVID-19 with objective to help alleviate these symptoms or as a possible solution. Specifically, by using T lymphocytes as living drugs equipped with a CAR construct directed against TH2 or viral epitopes. We hope to curb the severe complications and the poor outcomes (mortalities) associated with this pathology.
SARS-CoV-2, CAR T, Cytokine storm, Anti-CAR, TH2, Viral epitopes, Poor outcomes
Coronaviruses in the last two decades have caused serious infection and mortality in humans [1,2].
SARS-CoV-2, is a beta-coronavirus, similar to two other coronaviruses causing deadly infections [3]. But differences in sequences in the spike protein in SARS-CoV-2 results in its enhanced binding to angiotensin-converting enzyme 2 (ACE2) in human lung cells [4].
This zoonotic disease, COVID-19 not only affected people's physical health, but also destroyed mental health and floundered economic growth [5,6].
Globally, as of February 21, 2023, there have been 757,264,511 confirmed cases of COVID-19, including 6,850,594 deaths, reported to WHO and a total of 13,222,459,780 vaccine doses have been administered [7].
Communication has been critical during this pandemic, both for general guidelines and for new scientific information; however, there is a fine line between scientifically accurate information and the brevity and simplicity with which it should/could be communicated; sometimes trading accuracy for simplicity may result in a message that is more confusing rather than clarifying [8].
Interestingly, there is the principle of the iceberg in COVID 19, with symptomatic patients consisting the visible part of the iceberg.
Different countries exhibit varying results and this could be due to poor diagnostic tests, limited access to diagnostic tests to identify affected people in different countries and different percentages of asymptomatic people with COVID-19 in various communities.
Hence the interaction between these groups of people could increase the creation of new recombinant viruses which could be more harmful and more difficult to treat. This possible interaction has been demonstrated in a uniquely closed environment [9].
Also, this interaction can be seen by the variants of concern (variants that have been associated with an increase in the transmission or mortality of COVID-19) with the latest being variant B.1.1.529, commonly known as Omicron [7,10-12].
There are reports that vaccinated people made up 42% of fatalities in January and February during omicron. That’s compared with 23% at the peak of the delta wave [13]. This could be testament to this viewpoint in our opinion. However recent data suggests that people who are vaccinated are 20 times less likely to be hospitalized [13].
We strongly believe that primary prevention would have been the best option in an ideal situation but we are far from been in one; due to socio-economic difficulties (which could explain the disparity in vaccinations between countries) and the enormous human interaction given that the world is now a global village (Figure 1).
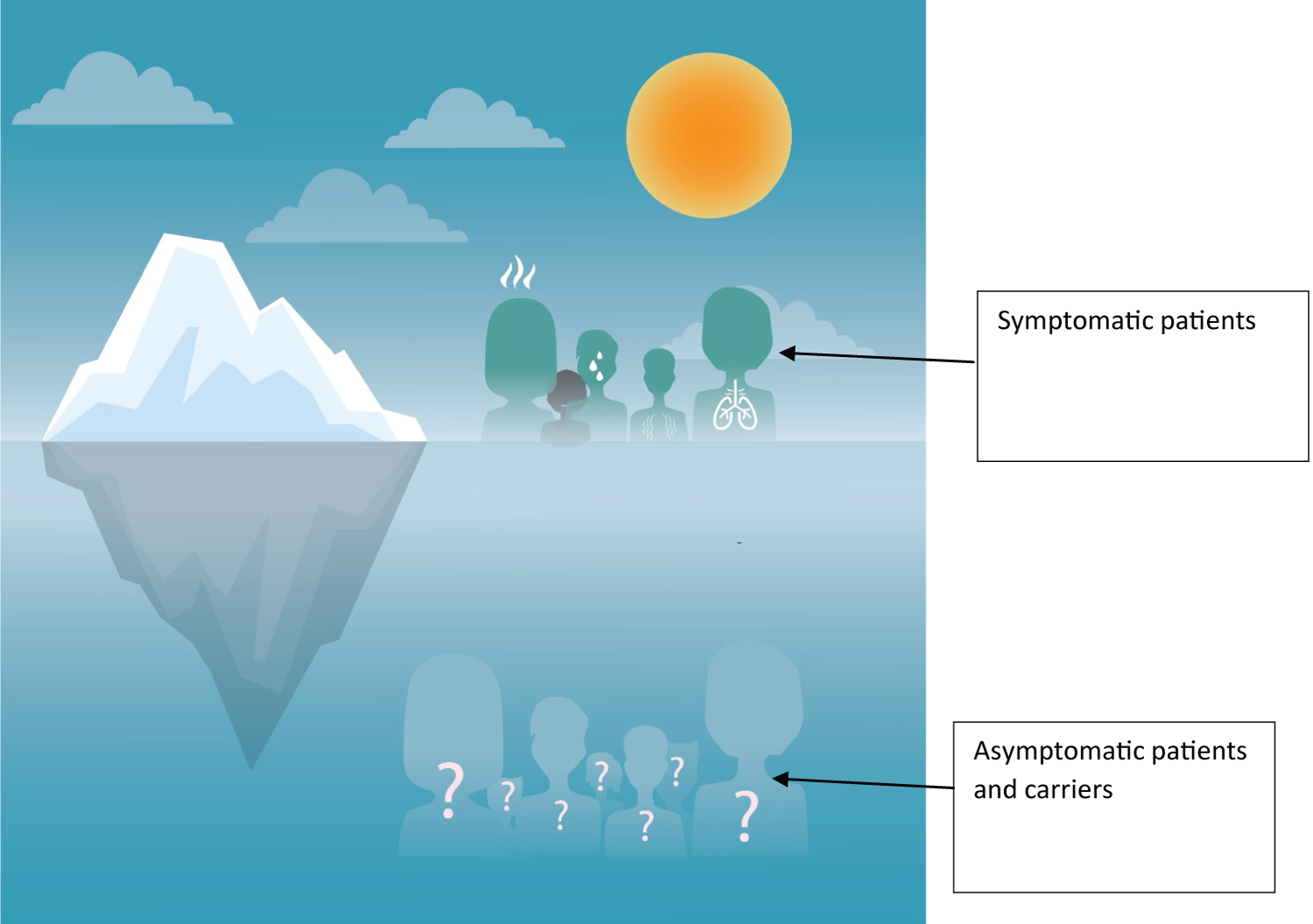 Figure 1: Iceberg principle in COVID-19.
View Figure 1
Figure 1: Iceberg principle in COVID-19.
View Figure 1
SARS-CoV-2 invades the host by virtue of angiotensin-converting enzyme 2 (ACE2) receptors broadly distributed on various tissues and immune cells [1,14]. The virus can cause a wide array of clinical manifestations ranging from mild to severe forms with fatal outcomes [1].
Evidence has demonstrated that deterioration of COVID-19 may be due immunopathological damage.
Particularly, separate studies have reported that highly elevated levels of pro-inflammatory cytokines are produced during the crosstalk between epithelial cells and immune cells in COVID-19, which has linked the cytokine storm (CS) with the severe complications and poor outcomes in this infection [1].
Due to their cytotoxic capacity, T cells emerged as attractive candidates for specific immunotherapy of cancer. A promising approach is the genetic modification of T cells with chimeric antigen receptors (CARs) [15].
The CAR-mediated recognition induces cytokine production and tumor-directed cytotoxicity of T cells [15].
Chimeric antigen receptor (CAR) T cell therapy represents a paradigm shift in the management of pediatric B-cell acute lymphoblastic leukemia (ALL) and adult B-cell non-Hodgkin lymphomas (NHL) [16].
There has been enormous progress in genetic engineering in the last decades paving the way for the development of CAR T cells.
CARs typically comprise an extracellular antigen recognition moiety fused via a flexible hinge and transmembrane region to an intracellular signaling unit, thereby combining the virtues of antibodies (high antigen-binding specificity) and immune cells (potent anti-tumor effector mechanisms) within one single fusion molecule [1].
Unfortunately, severe treatment-associated toxicities still restrain the widespread application of this promising technology. The most frequent side effects following CAR T-cell administration include cytokine release syndrome [4-7].
Given that organ injury in COVID 19 could be due to direct viral infection and immune overactivation, we intend to prevent these by using anti-CAR T engineered T cells.
Cytokine storm is the most life-threatening complication associated with COVID-19, characterized by a hyperactivated and proinflammatory state of the immune system [17-19]. Highlighting the importance of the immune system in the physiopathology of this disease [19-21].
Sufficient evidence has revealed the components and characteristics of CS in the patients with severe COVID-19, which are composed of an array of cytokines [1].
The initiation of COVID-CS induction during infection and the predominant causative cytokine in COVID-19 immunopathology remain largely unknown. Despite the lack of definite pathogen associated molecule pattern (PAMP) of SARS-CoV-2, in analogy with SARS-CoV and MERS-CoV, it can be speculated that upon cellular entry of SARS-CoV-2 via its ACE2 receptor, viral genomic single-stranded RNA or other RNA compositions (double-stranded RNA) as PAMPs can be sensed by the related pattern recognition receptors (PRRs), in host cells [1].
However, the protective IFN-I response is quickly and selectively abrogated by SARS-CoV-2 via different mechanisms. This is accompanied by an overwhelming production of pro-inflammatory cytokines in the context of COVID-19, which not only impairs viral clearance but also promotes paradoxical hyperinflammation including CS. Therefore, from the immunology perspective, COVID-CS may be an unfortunate event whereby the intended host immune response combating the SARS-CoV-2 has lost control and transformed into an inflammatory type.
In SARS-CoV-2 infection, the virus infects the respiratory epithelial tissue and activates local innate immune cells to release inflammatory cytokines and other chemokines, which then recruit more innate immune cells and activate adaptive immune cells (CD4+ and CD8+ T cells) from the peripheral tissues to produce sustained inflammatory which induce myelopoiesis and emergency granulopoiesis that further aggravate lung and epithelial damage.
In addition, overproduction of systemic cytokines gives rise to anemia through erythro-phagocytosis and macrophage activation as well as causes perturbation of coagulation and vascular hemostasis, resulting in capillary leak syndrome, thrombosis, and DIC. These events together lead to ARDS, multiorgan failure, and death.
It is worth noting that, the host immunoregulatory system is usually capable of retaining and fine-tuning the protective inflammation to an appropriate level. Regulatory cells such as Tregs, can produce regulatory cytokines like IL-10 and tumor growth factor-β to antagonize overactivated immune responses. However, aggressive inflammatory conditions such as CS cannot be calmed by the regulatory system’s natural ability.
COVID-CS is a complicated and dynamic inflammatory process caused by a group of cytokines from initiation, immune cell hyperactivation, to organ dysfunction. The development of precise therapeutic intervention in appropriate time is required to effectively control COVID-CS. In principle, the treatment strategy is to control ongoing inflammatory response by specifically or nonspecifically targeting inflammatory cytokines or related signaling pathways and to resume the host immunoregulatory system [1].
Excessive local release of cytokines is considered to be the determinant of pathological alterations and the clinical manifestation of ARDS. Overall, the primary pathological manifestations in the lung tissue are viral cytopathic-like changes, infiltration of inflammatory cells, and the presence of viral particles. Thus, severe lung injury in COVID-19 patients is considered as the result of both direct viral infection and immune overactivation [22].
Clinical research on adoptive transfer of T lymphocytes for infectious and malignant disorders is quite active. The use of cells transduced with T cell receptors (TCRs), in which tumor antigen recognition occurs through presentation on cell surface human leukocyte antigens (HLA), and the use of chimeric antigen receptors (CARs), which are typically specified by a single-chain variable region domain of an antibody and directed to a cell surface tumor-associated antigen, are fundamental approaches to live T cell immune therapy [23].
Chimeric antigen receptor T cells (CAR T) are a promising type of immunotherapy that uses genetically modified T cells to target cancer cells. Several CAR T cell products have recently demonstrated impressive efficacy against previously difficult-to-treat cancers. Additionally, second-generation CARs include a CD28 or 4-1BB co-stimulatory endodomain [24].
Over the last few decades, there has been tremendous progress in genetic engineering, paving the way for the development of CAR T cells.
CARs typically consist of an extracellular antigen recognition moiety fused to an intracellular signaling unit via a flexible hinge and transmembrane region, combining the benefits of antibodies (high antigen-binding specificity) and immune cells (potent anti-tumor effector mechanisms) within a single fusion molecule [1].
Immunotherapies have showed promise, but they are far from flawless as evidenced with certain serious treatment-associated toxicities.
Unlike conventional cytotoxic chemotherapy or small molecule inhibitors, immunotherapies have specific toxicities that might be fatal, such as cytokine release syndrome (CRS) and CAR-related encephalopathy syndrome (CRES) [24]. With cytokine release syndrome being the most common side event after CAR T-cell administration [4-7].
Several strategies have been tested to counteract these effects, from non-specific immunosuppression to selective CAR-engineered T cell ablation.
The latter technique is currently under huge research and is based on the transgenic introduction of either suicide genes or elimination marker genes.
Following the introduction of these therapeutic agents, selective CAR T-cell depletion is achieved through complement-dependent cytotoxicity (CDC) and antibody-dependent cellular cytotoxicity (ADCC).
However, each method has inherent limits that could prevent it from having a wide range of therapeutic applications. These procedures also have serious toxicities. These include immunogenicity [20] the proposed depletion marker's enormous size (more than 130 amino acids), the reliance on the patients' immune systems (ADCC, CDC), and the occurrence of on-target adverse effects brought on by mAb (monoclonal antibodies) recognizing healthy tissue [21].
The term CRS (cytokine release syndrome) here refers to the excessive release of cytokines (IL-1, IL-6, IFN-, and IL-10) by CAR-modified immune cells or bystander innate immune cells (macrophages, monocytes, dendritic cells, and other immune cells). Excess cytokines can result in vascular leakage, respiratory failure, coagulopathy, and multi-organ system dysfunction.
The production of a significant amount of interferon gamma (IFN-) and/or tumor necrosis factor alpha (TNF-) by CAR-T activated cells, which in turn activates macrophages, dendritic cells, and endothelial cells, is one possible cause causing CRS. These cells can release more proinflammatory cytokines after being stimulated by IFN- or TNF [8].
This genetically modified T cells that constitute the CAR T cells can induce a cytokine storm.
Control and reversal of toxicity have emerged as crucial components of CAR T-cell treatment due to the extremely long-term survival and proliferation capacity of genetically altered T cells [25].
Bachmann, et al. suggested integrating a particular peptide epitope (E7B6) into the CAR architecture that can be utilized as an intrinsic elimination tag as a way to address this. This E-tag into the CAR's extracellular spacer region should stop CAR T-cell escape, reducing the therapy's cytotoxic effects while also increasing its safety [25].
Therefore, for selective and rigorous CAR T-cell elimination, Bachmann, et al. used a short peptide epitope (E-tag) directly inserted into the CAR architecture [25-27].
Additionally, Bachmann, et al. sought to avoid reliance on pharmacological drugs whose therapeutic effect invariably declines due to their short half-life, which is a significant drawback, by using T lymphocytes as living drugs that are equipped with a CAR construct directed against a targetable portion of the therapeutic CAR [25].
With the other experiments, the cloning and structural characteristics of conventional CARs as well as the universal chimeric antigen receptor (UniCAR) 28/ζ construct with the incorporation of the peptide epitope E7B6 (E-tag) as a targetable moiety in the extracellular spacer region are described in detail [25,27-29].
In the hinge region of CARs, a peptide tag called E7B6 is recognized by the αE-tag CAR construct. When an antigen is recognized, these αE-tag CAR effector cells cross-link with target cells, which should cause the elimination of the latter.
Furthermore, Bachman, et al. demonstrated that the cytotoxic effect was caused by the incorporated peptide's specific recognition and binding (αE-tag CAR).
The anti-CAR therapy has been demonstrated to prevent the cytotoxic effects due to CAR T therapy and has been described in detail by Bachmann, et al. [25].
Our hypothesis could act by targeting impaired viral clearance and/or lymphocytes.
Against viral infections, the adaptive immune system, particularly T cells, plays an important role [29]. T cell responses can be divided into two types: effector cytotoxic cells (CTLs) and T helper cells (Th) [19,30,31].
Data suggest that cytokine storms may contribute to the pathophysiology of severe COVID-19 disease together with reduced Th1 antiviral adaptive responses [32].
Adults with COVID-19 experience acute peripheral T cell depletion, the degree of which is positively correlated with the severity of the disease, whereas asymptomatic patients and children typically retain peripheral T cell numbers [33].
The progression of the disease in COVID-19 has been associated with the Th1/Th2 balance. An appropriate Th1 immune response, once a viral infection has been recognized, can eradicate it. The cytokine storm that results from an overactive immune response and increased cytokine production, however, leads to a Th2-type response and a poor prognosis for COVID-19 [33-35].
In the same light, research has demonstrated that elevated IL-15 levels and a strong Th2 response are linked to the disease's deadly prognosis and that senescent Th2 cell percentage was an independent risk factor for death [19].
Also, COVID-19 patients had significantly fewer Th1 and more highly activated Th2 cells than the reference population [19].
Th1 coordinated immune response during SARS-CoV-2 infection has been associated with a good prognosis and resolution of COVID-19. The type of Th response influenced disease outcome, as 78% of patients who died had an overreactive Th2 response [19,36,37].
Because their cytokines stimulate antibody production, Th2 hyperactivation could explain why severe patients have higher antibody titers than mild or asymptomatic patients [19,36,37].
Moreover, within the cell, M. Bacillus Calmette-Guerin (BCG) vaccination stimulates a Th1 response, guarding against the development of tuberculosis in humans. With incredibly unexpected results, the effectiveness of the BCG vaccine has been researched in COVID-19 patients. Patients who received the vaccine and developed Th1 responses had lower death and infection rates [38]. A recent research, has emphasized the significance of Th1 hypoactivation and Th2 overreaction, followed by exhaustion, as they are related to a worse prognosis [19].
Hence by directing genetically modified anti-CAR T cells against these lymphocytes (Th2) we hypothesize that this could limit or prevent the cytokine storm.
Also, could we direct these genetically modified anti-CAR T cells against the viral epitopes to help limit or prevent this cytokine storm.
The SARS-CoV-2 virus is a single-stranded enveloped positive-sense RNA virus that belongs to the β-coronavirus [39,40].
The proteins encoded by the genome of SARS-CoV-2 are comprised of structural (SPs) and nonstructural proteins (NSPs), as well as accessory proteins [41,42]. SPs mainly include spike (S), membrane (M), envelope (E), and nucleocapsid (N) proteins [39,43].
The virus receptor-binding domain (RBD) contains several antigenic epitopes. Those antigenic epitopes, also known as antigenic determinants, are the binding sites of host antibodies. The antigenic epitope plays an important role in activating the host CD4 and CD8 T cell immune response [44].
Therefore, the S protein, the RBD domain, and antigenic epitopes pave the road for the therapeutic strategies [39].
So by targeting these viral epitopes especially of the S protein, we might to control the interactions and underlying mechanisms between the host and virus infection. More so, research has shown that SARS-CoV-2 uses the structural function of the S protein to weaken or escape host immune surveillance [39].
Hence by creating a genetically modified CAR T antagonist, could help curb the cytokine storm, given that thus aggressive inflammatory condition (the cytokine storm) cannot be calmed by the regulatory system’s natural ability and also given the non-precise nature of general immunosuppression.
We propose an antagonist for CAR-T (Chimeric Receptor Antigen T cell) therapy as a solution to prevent cytokine storm in patients with COVID-19 and hence reduce the morbidity and mortality due to COVID 19.
The SARS-CoV-2 virus can cause a wide range of manifestations from mild to severe/critical forms with fatal outcomes; fatal outcomes with increased morbidities and mortalities which we aim to prevent.
Cytokine storm during the COVID 19 infection is fatal to the patients. This unfortunate event causes deaths due to ARDS and multi-organ failure.
CAR-T therapies have as one of its effects inducing a cytokine storm. So, by using immunopharmacology more precisely, a genetically modified antagonist of this therapy; we intend to prevent or curb the adverse effects caused by this cytokine storm which can be helpful in severe SARS-CoV-2 to reduce the mortalities and morbidities associated with this pandemic (Figure 2).
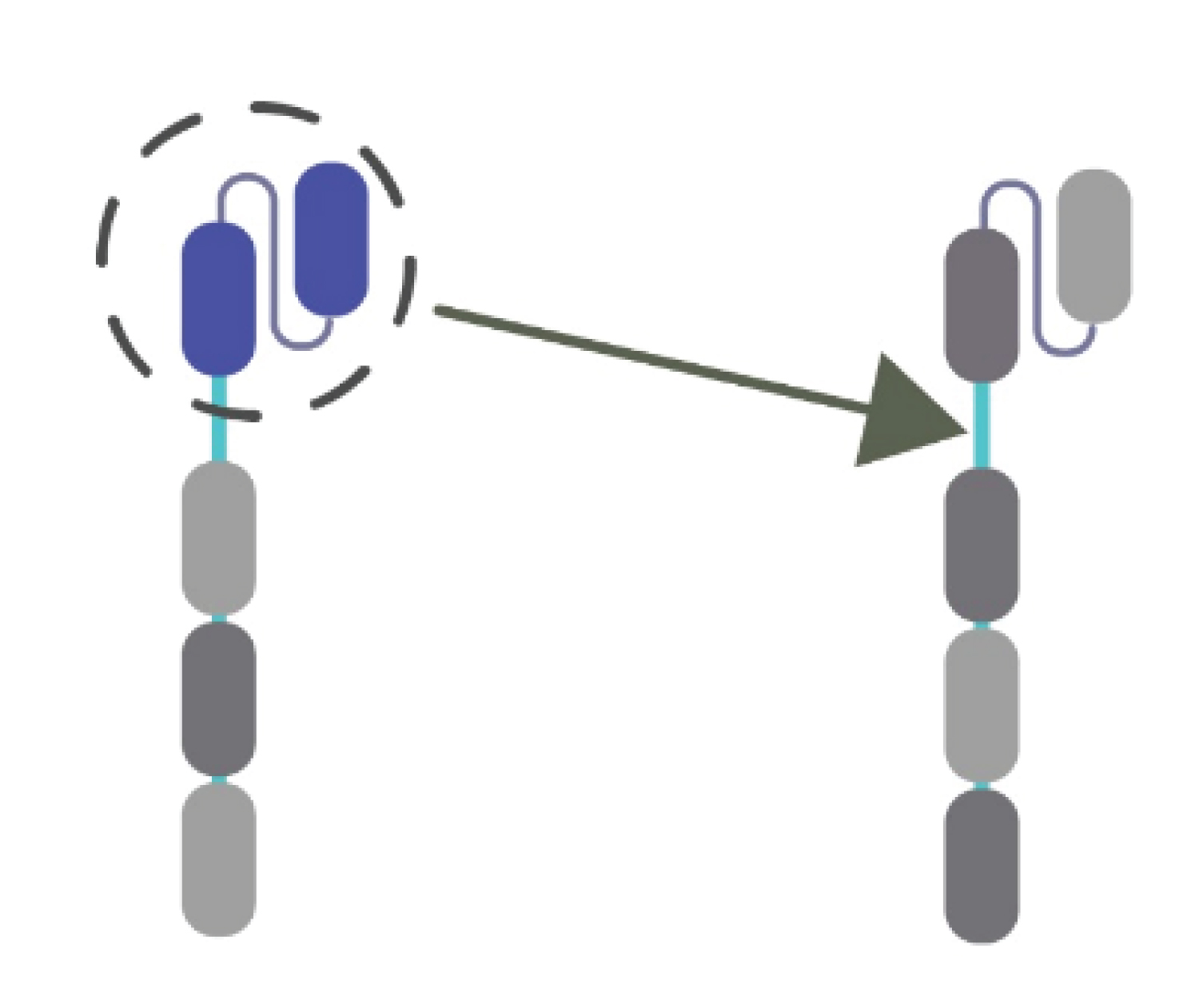 Figure 2: Target; CAR T cells.
View Figure 2
Figure 2: Target; CAR T cells.
View Figure 2
Anti-CAR targets selectively and precisely eliminates CAR T-cell with the help of its short peptide epitope (Figure 3).
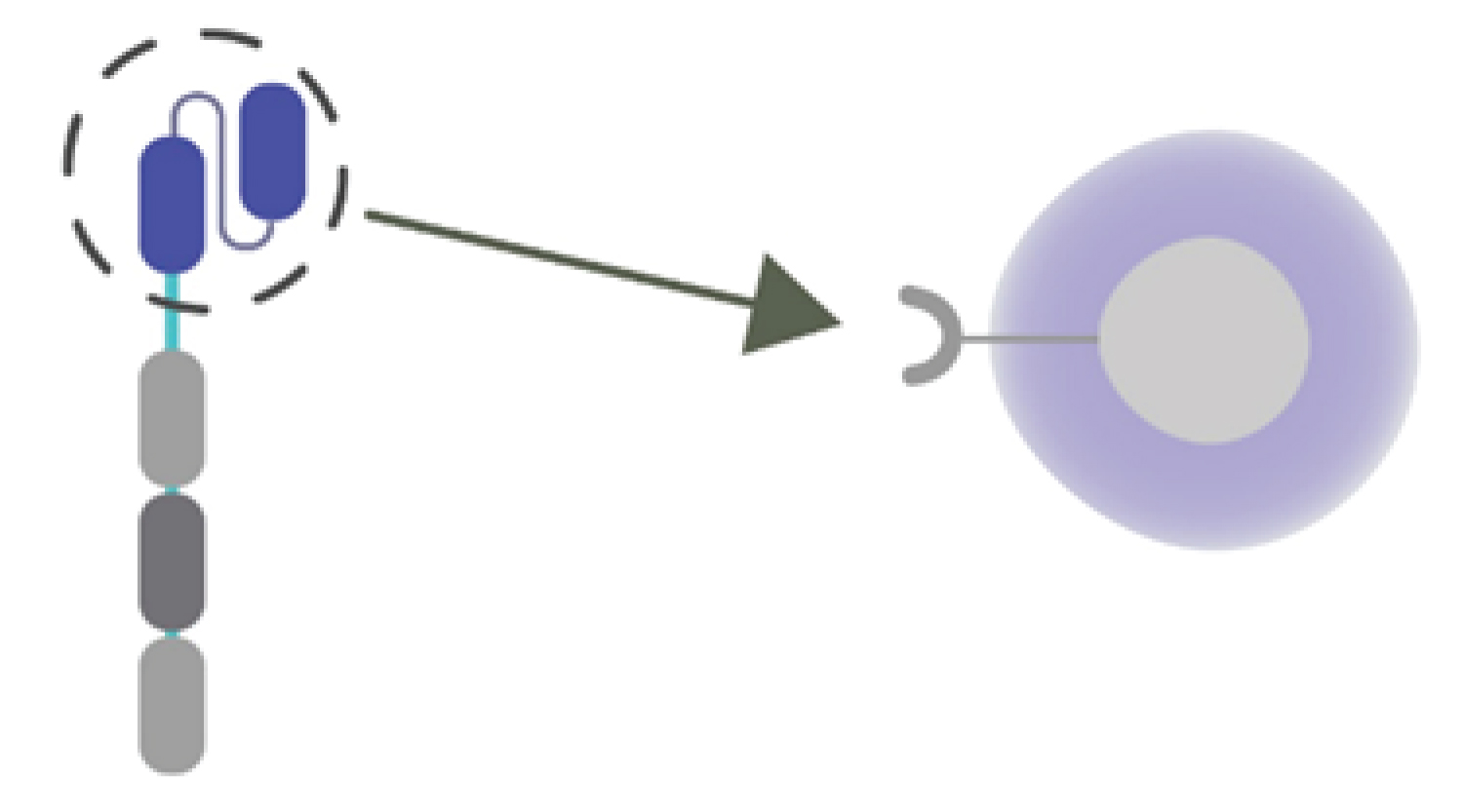 Figure 3: Target; Th2 lymphocytes or viral epitopes.
View Figure 3
Figure 3: Target; Th2 lymphocytes or viral epitopes.
View Figure 3
We propose using a genetically modified anti-CAR T cells against Th2 lymphocytes to prevent cytokine storm or viral epitopes.