Most studies on cadmium toxicity focus on its singly effect neglecting the fact that it does not occur alone in the environment but coexist along with other metals. The current study was therefore designed to examine the possible influence of iron (Fe) on cadmium (Cd) toxicity in the gonad of female rats. Twenty adult female albino rats used in this study were divided into four groups of five rats each. The groups were designated as group A-control (rats exposed to exogenous Cd-free diet), group B (rats exposed to Cd-tainted diet), group C (rats exposed to Fe-tainted diet) and group D (rats exposed to combined Cd and Fe- tainted diet). The rats were exposed to Cd (0.229 mg/L) and Fe (1.900 mg/L) through food chain-mediated diet by feeding them for a period of 4 weeks. Results show that, there was significant decrease (p ≤ 0.05) in serum catalase (CAT) activities of rat exposed to Fe only and Cd+Fe relative to the control but a significant increase (p ≤ 0.05) in ovary CAT activity of rats exposed to Cd+Fe. The malondialdehyde (MDA) level in ovary of rats exposed to Fe only was significantly decreased (p ≤ 0.05) when compared to the control. Exposure of rats to Cd only, Fe only and to the combination of Cd and Fe significantly decreased (p ≤ 0.05) the cholesterol level in the ovary when compared to the control. The levels of Luteinizing Hormone (LH), Follicle Stimulating Hormone (FSH) and estradiol in sera of rats exposed were significantly increased (p ≤ 0.05) in Cd only and in combined Cd+Fe groups when compared to the control but were significantly decreased in the Fe only exposed group. The level of testosterone was significantly increased (p ≤ 0.05) in combined Cd+Fe group relative to the control. Histological study revealed discohesive follicular cells and few primary follicles in ovary of rats exposed to Cd only and Cd+Fe. The results from this study suggest that cadmium only as well as its combination with iron caused the biochemical changes induced in the ovary. Since Cd only was able to cause the changes observed, it can therefore be suggested that iron was unable to ameliorate the effect of this changes. Findings from histological examination of the ovaries show that there was profound disintegration with follicular damage appear to corroborate the findings made in this study. In conclusion, cadmium toxicity is apparently enhanced in the presence of iron as evidence by its effects on cadmium-associated alteration in the serum levels of estradiol and testosterone of rats exposed.
Cadmium, Iron, Diet, Gonads, Hormone
One environmental pollutant which has been shown to be highly toxic to both humans and animals is cadmium [1,2]. Cadmium accumulation in the environment is due to its widely spread industrial use. If gained entrance into the body, it can accumulate in the blood, kidney, and liver as well as in the reproductive organs [1-3]. Cadmium has been shown to target multiple organs following acute intoxication, causing nephrotoxicity, immunotoxicity, osteotoxicity and reproductive toxicity [4]. Cadmium is known to affect reproductive organs [5,6].
Cadmium does not occur alone but exists along with other pollutants in water, land and even in air. Any organism exposed to it will suffer the combine toxic effects of these pollutants. Cadmium (Cd) present in the air, drinking water and food has the potential to affect the health of people, mainly those who live in highly industrialized regions due to their industrial activities [7]. Cd affects placental function [7] and even gonadal function [8]. Cd toxicity is influenced by the presence of other heavy metals such as iron, lead, zinc, copper, manganese, chromium and nickel [9].
However, since cadmium does not occur alone, it is therefore better to look at its combine toxic effect rather than its single effect. The purpose of the present study was therefore aim to evaluate the effect of cadmium (Cd) and iron (Fe) toxicity singly and combined on the reproductive organs (gonads) of female rats, as a model for human representative. The rats were exposed to Cd and Fe tainted diet through feeding for 4 weeks. The study investigates the gonadal effects of food chain- mediated exposure of rats to cadmium and iron.
Wistar rats: Twenty adult female albino rats were obtained from Animal Science Department, University of Ibadan, Oyo State. The animals were acclimatized for a period of one week before the commencement of the treatments.
Catfish: Forty post-juvenile catfish (source of protein in diet) used for this study were obtained from the Department of Animal Science and Environmental Biology, Adekunle Ajasin University, Akungba Akoko, Ondo State. The fish were acclimatized for a period of two weeks before the commencement of the treatment.
Chemicals/Reagents: All chemicals used for this study were of analytical grade. Cadmium chloride hemidihydrate (CdCl2.2.5H2O) was the product of Kermel, Germany. Iron (II) chloride tetrahydrate (FeCl2.4H2O) was the product of JHD, China. Total cholesterol assay kit was the product of Randox, United Kingdom. ELISA kits were product of Monobind Inc, USA.
Preparation of stock cadmium/iron solution and cadmium/iron tainted water: Stock solution of cadmium/iron and cadmium/iron tainted water were prepared as described by Ogunbiyi, et al. [10].
Treatment of catfish: The catfish were assigned into four experimental groups of 5 fish each after the two weeks of acclimatization. The fish were fed twice daily with vital fish feed (Livinco fish feed Ltd. Ikare, Ondo State) during the period of the study and the actual quantity of feed used was described by Obi, et al. [11].
Group A fish (control) were maintained in water that had no exogenous addition of either cadmium or iron. Group B fish were maintained in water tainted with exogenous cadmium (0.229 mg/L) only. Group C fish were maintained in water tainted with exogenous iron (1.900 mg/L) only. Group D fish were maintained in water tainted with both cadmium (0.229 mg/L) and iron (1.900 mg/L) simultaneously.
Each group was kept in 25 litres of water in 40-litre capacity plastic buckets with perforated lids and housed in a room with reduced illumination (temperature 25 ± 2 °C). The treatments were done for twelve weeks to achieve satisfactory bio-accumulation of metals in the fish. At the end of twelve weeks, each fish was rendered motionless by being "buried" in ice crystals for 5-10 minutes. They were sacrificed by hitting a stick on the head. The sacrificed fish were oven-dried at 110 °C for 24-hour, milled and used as source of protein in the formulated diets.
Formulation of tainted diet: The tainted diet was compounded following the method described by Asagba and Obi, et al. [12].
Treatment of rats: The rats were divided into 4 experimental groups of 4 rats each after one-week acclimatization period. Group A, the control rats were exposed to exogenous Cd-free diet. Group B rats were exposed to Cd-tainted diet. Group C rats were exposed to Fe-tainted diet and group D rats were exposed to combined Cd and Fe- tainted diet. The rats were maintained in a standard laboratory conditions using a well ventilated wooden cages with 12 h light: 12 h dark schedules at 27 ± 1°C and fed with standard rat pelleticized chow (Bendel Feed Flour Mill, Ewu, Edo State) prior to the study and drinking water ad libitum prior to and during the period of the study. The rats were exposed to Cd (0.229 mg/L) and Fe (1.900 mg/L) tainted diet through food chain-mediated exposure. The exposure period lasted for 4 weeks.
National Institute of Health (NIH) Guidelines for the Care and Use of Laboratory Animals [13] were strictly adhered into during the experiment.
At the end of four weeks of treatment, all rats were weighed and anaesthetized in anaesthetic ether saturated chamber and while under anaesthesia, the blood was collected by heart puncture and transferred to plain sample bottles. The ovary was placed on ice, weighed and subsequently homogenized using a pre-cooled mortar and pestle.
Preparation of blood serum and tissue homogenates: The collected blood samples, after clotting were centrifuged at 3,000 rpm for 15 minutes and the supernatant obtained (sera samples) were stored at minus 20°C until required for biochemical assays. The tissue (ovary) was chopped into very small pieces and homogenized in ice-cold physiological saline (0.9%) to obtain 10% homogenates (1: 9 w/v). The resulting homogenates were centrifuged at 5,000 rpm for 15 minutes and the supernatants obtained stored at minus 20°C until required for biochemical assays.
Serum and ovary cholesterol were estimated based on the enzymatic endpoint method of Allain, et al. and Roeschlaw, et al. [14,15] as described in Randox cholesterol assay kit leaflet.
Malondialdehyde (MDA) level was estimated based on the amount of thiobarbituric acid reactive substances (TBARS) which is produced as an indicator of lipid peroxidation as described by Guttridge and Wilkings, et al. [16]. Values of TBARS are quantitated using a molar extinction coefficient of 1.56 x 105 M-1cm-1 and expressed in terms of malondialdehyde (MDA) units per gram tissue. The method of Misra and Fridovich [17] was followed in the determination of superoxide dismutase (SOD) activity. The SOD activity was computed and expressed as described by Baum and Scandalios [18] in which a unit represents the amount of the enzyme required for 50% inhibition of epinephrine in 1 min. Catalase (CAT) activity was determined by following the method described by Cohen, et al. [19]. This estimation is based on the measurement of the rate of decomposition of hydrogen peroxide (H2O2), after the addition of the hormogenate supernatant which was the source of the catalase.
The reproductive hormone levels were assayed in the sera samples and estimated based on the standard protocols of enzyme- linked immunosorbent assay (ELISA) of Braunstein, et al. [20], Winter and Faiman [21], Klopper and Fuchs [22] and Tietz [23] as described in ELISA kit leaflets for luteinizing hormone (LH), follicle stimulating hormone (FSH), estradiol and testosterone respectively.
Histological study on ovaries obtained from experimental rats was carried out following the method of Kiernan [24]. The ovaries were immersed in Bouin's fixative for 12-24 h after removal from the animals until required for histological examination. It was later dehydrated, embedded in paraffin, sectioned (5 μm), and stained with haematoxylin and eosin and viewed for analysis using Omax research microscope.
The experimental results were expressed as mean ± standard error of the means (SEM) and were subjected to one-way analysis of variance (ANOVA) obtained from Graphpad Prism 6.0. Significant levels were set at p ≤ 0.05 while Tukey's multiple range comparison tests were used to determine significant difference between groups.
The cholesterol level in serum and ovary of rats exposed to Cd and Fe are presented in Table 1.
Table 1: Cholesterol level in serum and ovary of rats after 4 weeks of exposure. View Table 1
Exposure of rats to cadmium and iron had no effect on the serum cholesterol level of rats when compared to control. There were significant decreases (p ≤ 0.05) in cholesterol level of ovary of rats exposed to cadmium and iron as separate entities as well as cadmium plus iron combined when compared to control.
The MDA levels, as well as SOD and CAT activities in the serum and ovary of rats at the end of 4 weeks exposure to Cd and Fe were presented in Table 2 and Table 3.
Table 2: Effects of Cd and Fe on MDA levels & antioxidant enzyme activities in the serum. View Table 2
Table 3: Effects of Cd and Fe on MDA levels & antioxidant enzyme activities in the ovary. View Table 3
There was no significant difference in MDA levels and SOD activities in serum of rats exposed to both metals when compared to the control whereas a significant decrease (p ≤ 0.05) in serum CAT activities of rats exposed to iron only and cadmium plus iron test diets was observed when compared to control but this was not statistically different from cadmium- test diet.
The MDA level was significantly decreased (p ≤ 0.05) in ovary of rats exposed to iron test diet when compared to the control. There was significant increase (p ≤ 0.05) in CAT activities in ovary of rats exposed to Cd plus Fe concurrently when compared to the control.
The level of reproductive hormone in serum of rats after 4 weeks exposure were presented in Figure 1, Figure 2, Figure 3 and Figure 4 respectively.
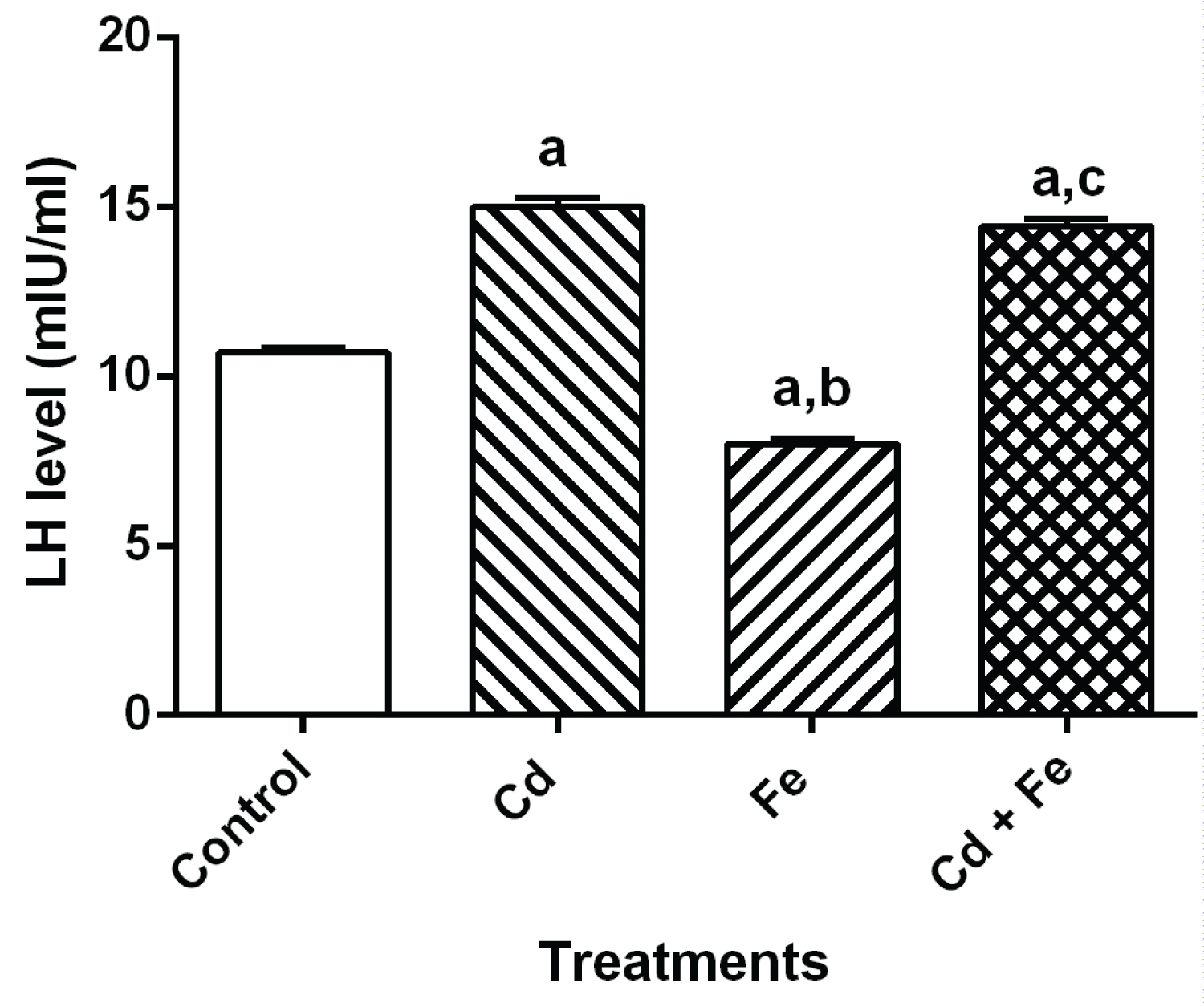 Figure 1: LH level in serum of rats. View Figure 1
Figure 1: LH level in serum of rats. View Figure 1
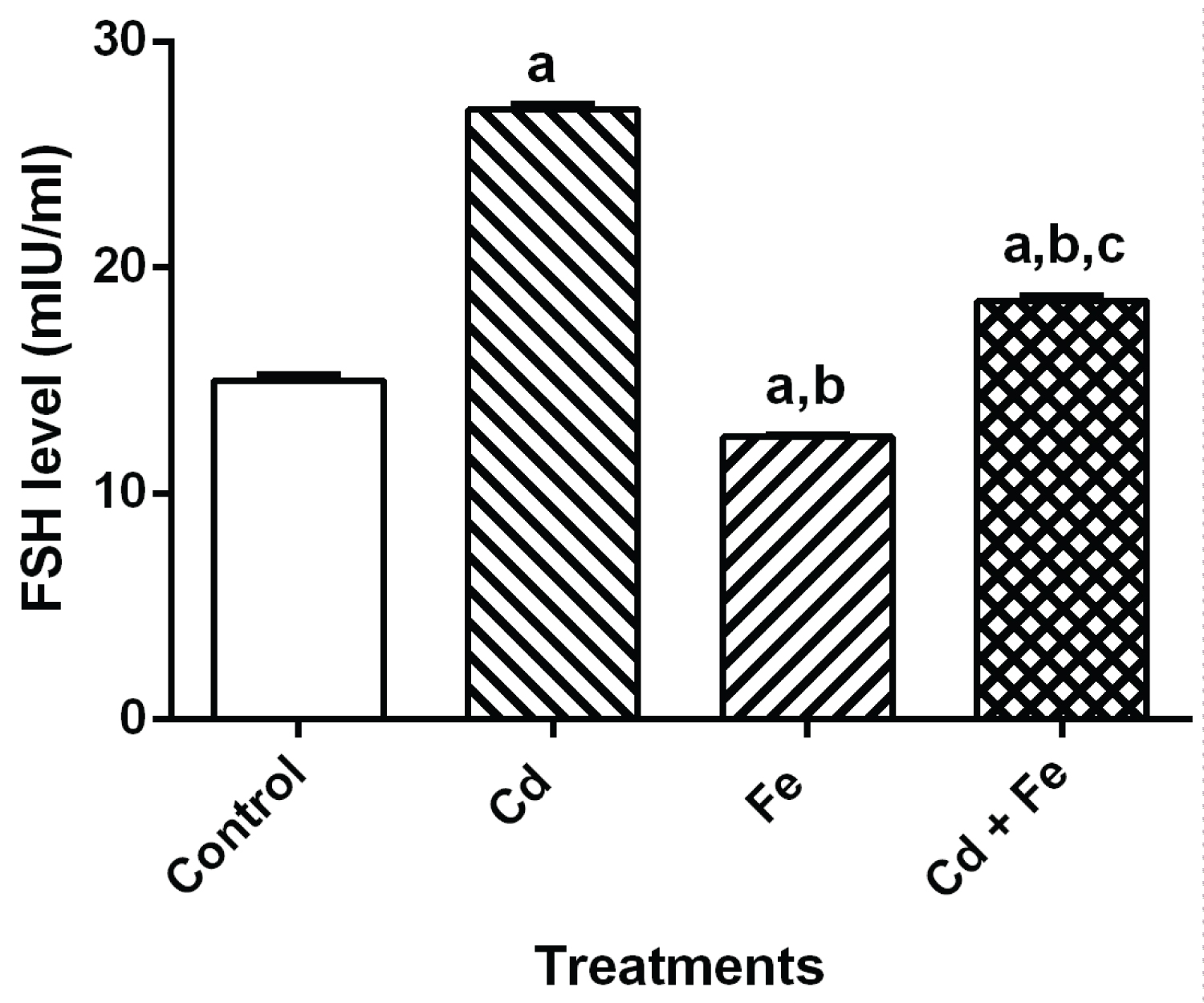 Figure 2: FSH level in serum of rats.
Figure 2: FSH level in serum of rats.
Values represent Mean ± SEM (n = 3). a Values with superscripts 'a, b and c' are significantly different (p ≤ 0.05) relative to the value of the control, the cadmium only and the iron only groups.
View Figure 2
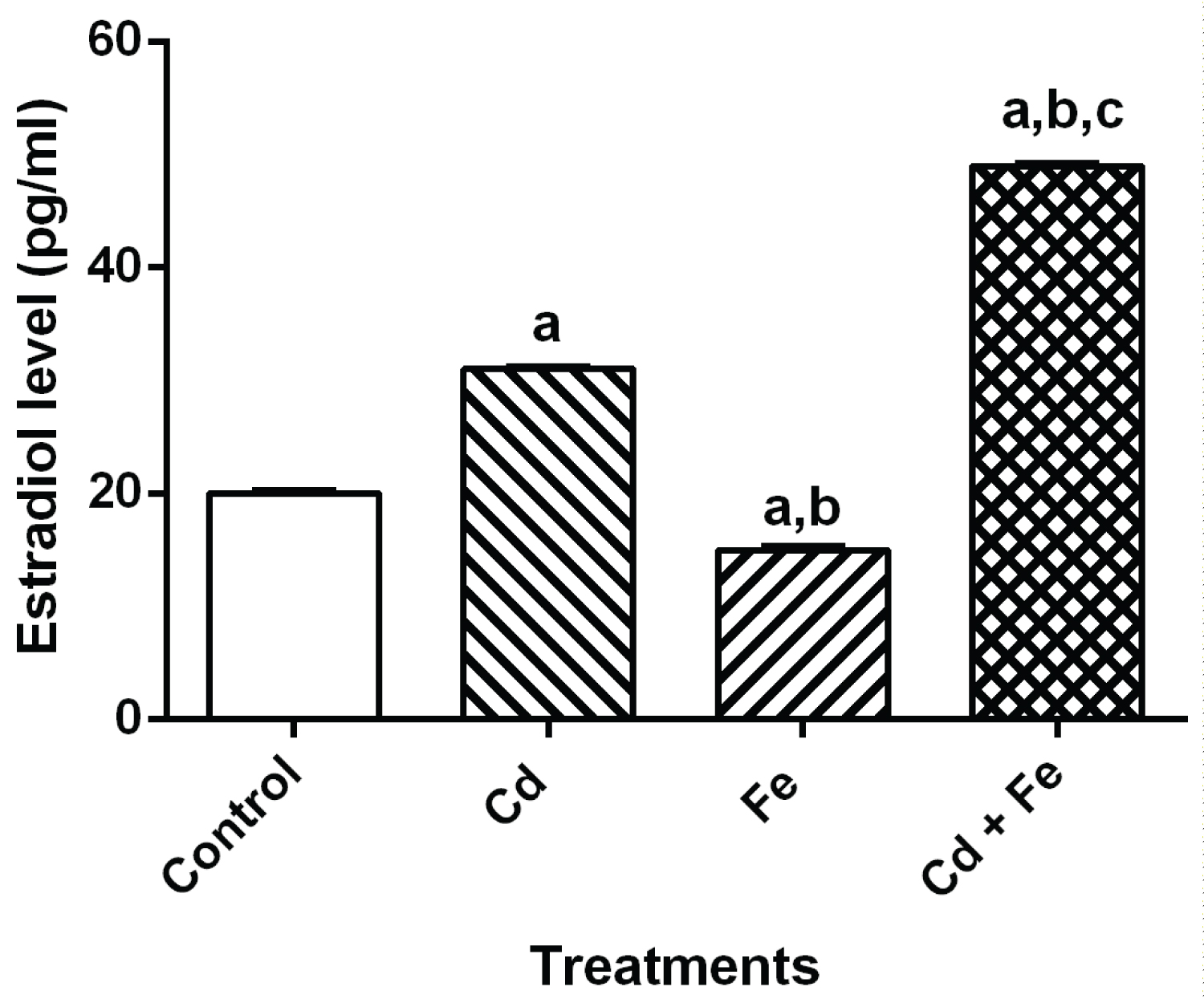 Figure 3: Estradiol level in serum of rats. View Figure 3
Figure 3: Estradiol level in serum of rats. View Figure 3
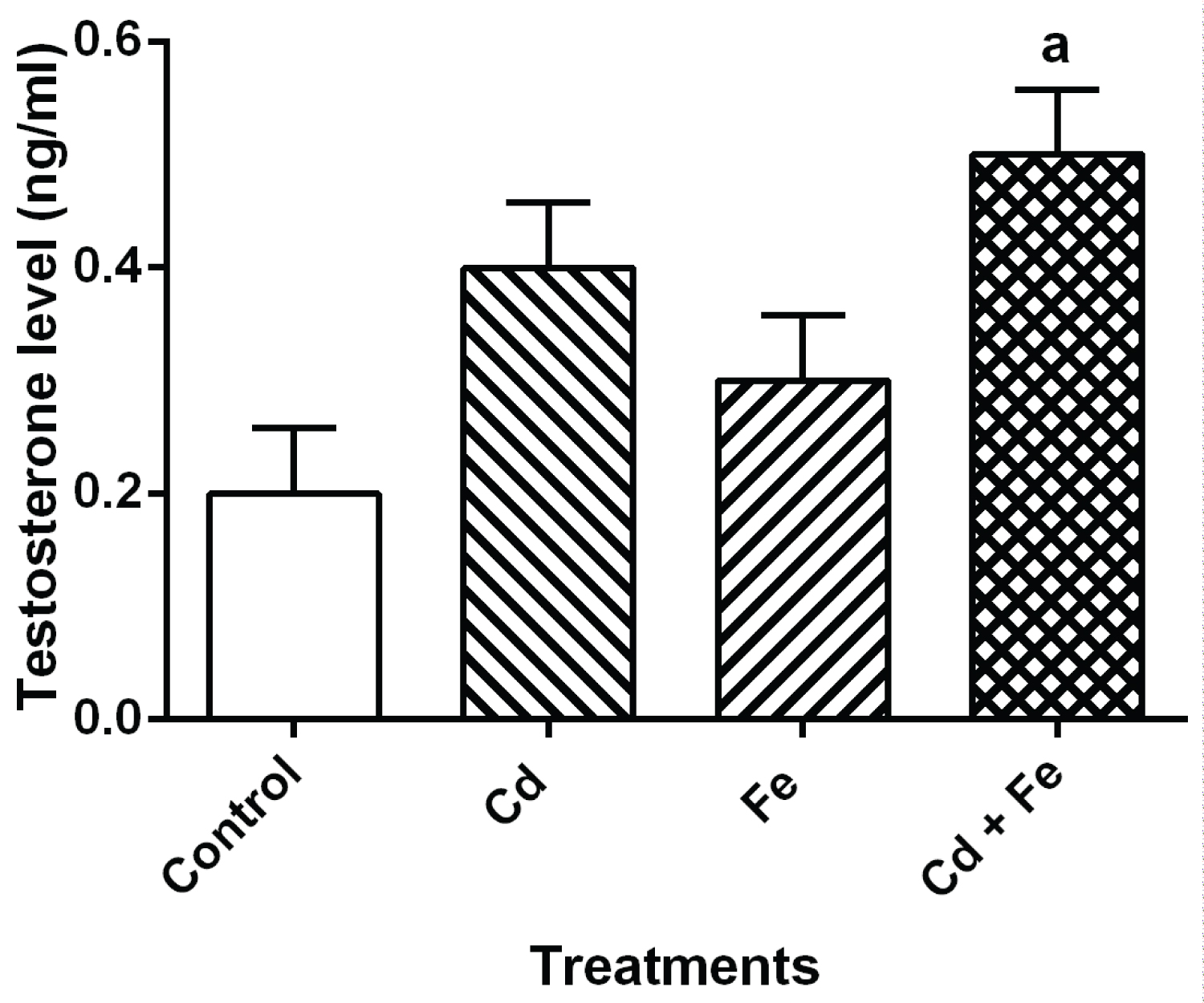 Figure 4: Testosterone level in serum of rats.
Figure 4: Testosterone level in serum of rats.
Values represent Mean ± SEM (n = 3). a Values with superscripts 'a, b and c' are significantly different (p ≤ 0.05) relative to the value of the control, the cadmium only and the iron only groups.
View Figure 4
Exposure of rats to both Cd and Fe resulted in increased levels of LH and FSH. When compared to control, there were significant increase (p ≤ 0.05) in both LH and FSH levels of rats exposed to cadmium and cadmium plus iron groups via food chain-mediated test diet but a significant decrease was observed in rats exposed to iron- tainted diet.
The levels of estradiol and testosterone were significantly increases when exposed to both Cd plus Fe. When compared to control, there was significant increase (p ≤ 0.05) in estradiol level of rats exposed to cadmium and cadmium plus iron tainted diet groups but a significant decrease in rats exposed to iron tainted diet group. Also, there was a significant increase (p ≤ 0.05) in the testosterone level of rats exposed to both metals concurrently.
Examination of the photomicrograph of section of ovary obtained from rat that were not exposed to Cd and Fe (Plate 1) revealed the presence of normal primary follicles with no visible lesion.
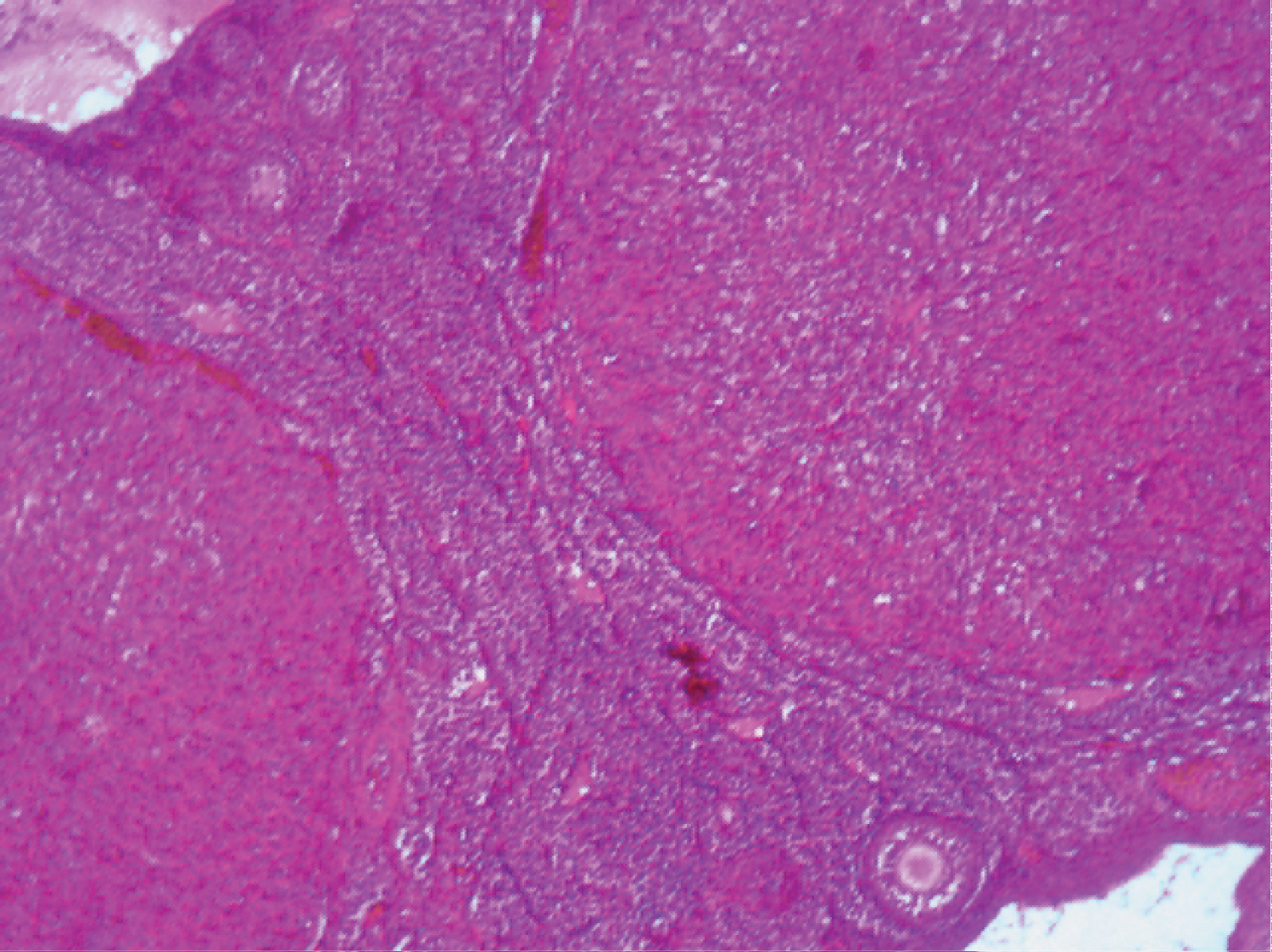 Plate 1: Photomicrograph of section of ovary from control rat (H and E x100). View Plate 1
Plate 1: Photomicrograph of section of ovary from control rat (H and E x100). View Plate 1
Examination of the photomicrograph of section of ovary obtained from rat exposed to Cd (Plate 2) revealed the presence of atretic follicles.
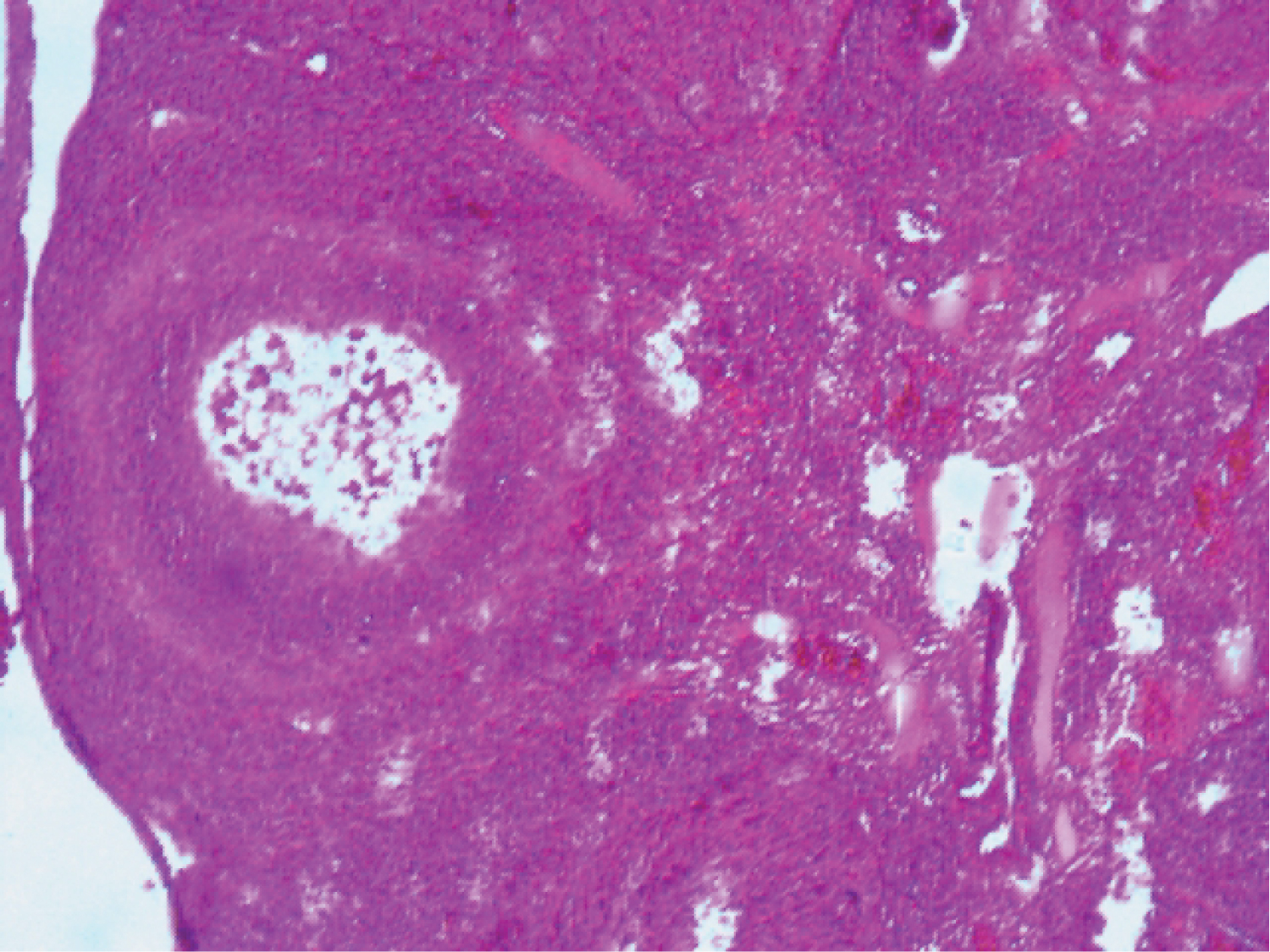 Plate 2: Photomicrograph of section of ovary from Cd exposed rat (H and E x100).
View Plate 2
Plate 2: Photomicrograph of section of ovary from Cd exposed rat (H and E x100).
View Plate 2
Examination of the photomicrograph of section of ovary obtained from rat exposed to Fe (Plate 3) revealed the presence of a mild distortion in the follicle.
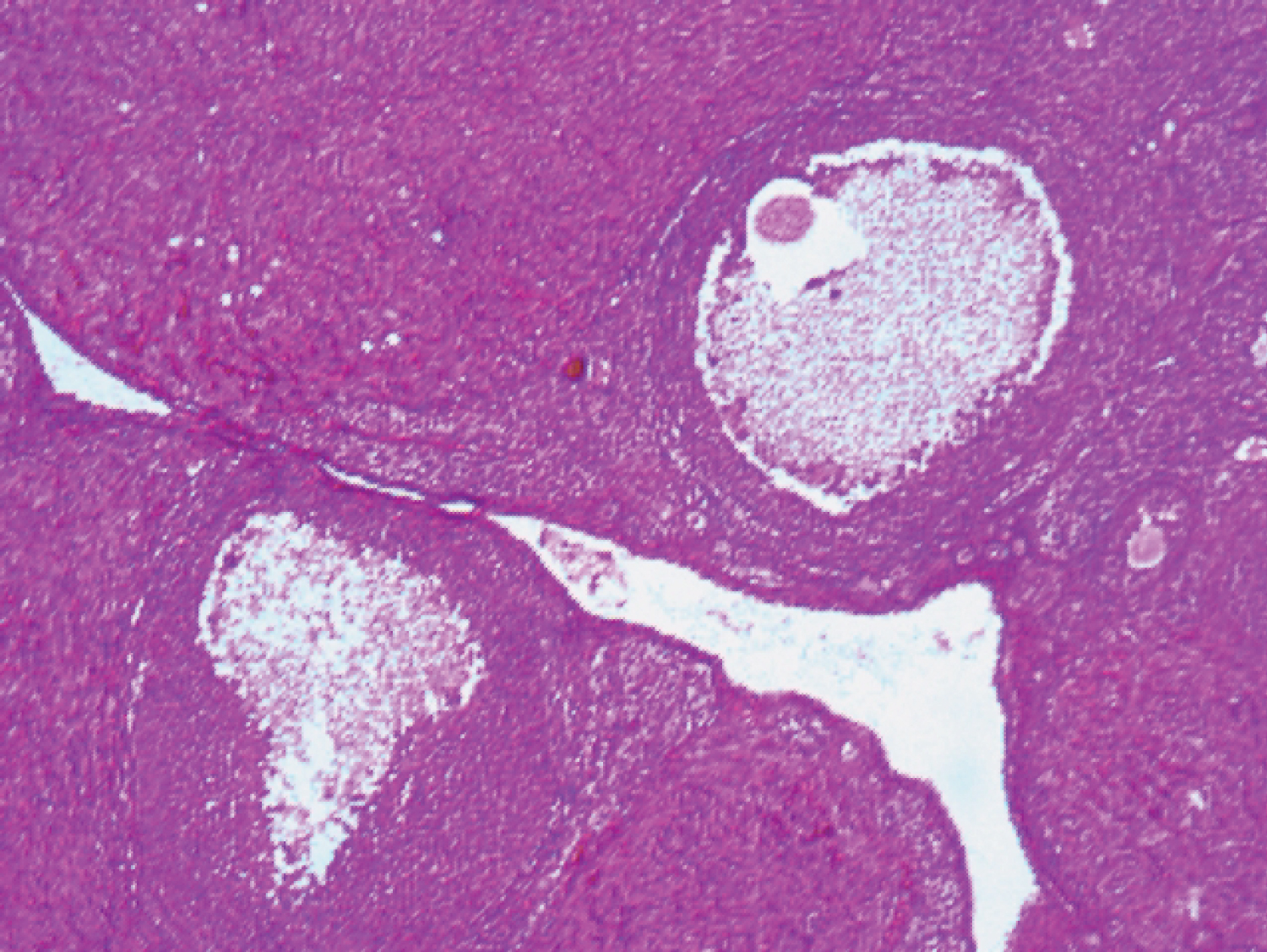 Plate 3: Photomicrograph of section of ovary from Fe exposed rat (H and E x100). View Plate 3
Plate 3: Photomicrograph of section of ovary from Fe exposed rat (H and E x100). View Plate 3
Examination of the photomicrograph of section of ovary obtained from rat exposed to Cd plus Fe (Plate 4) revealed the presence of gross follicle disintegration and few primary follicles.
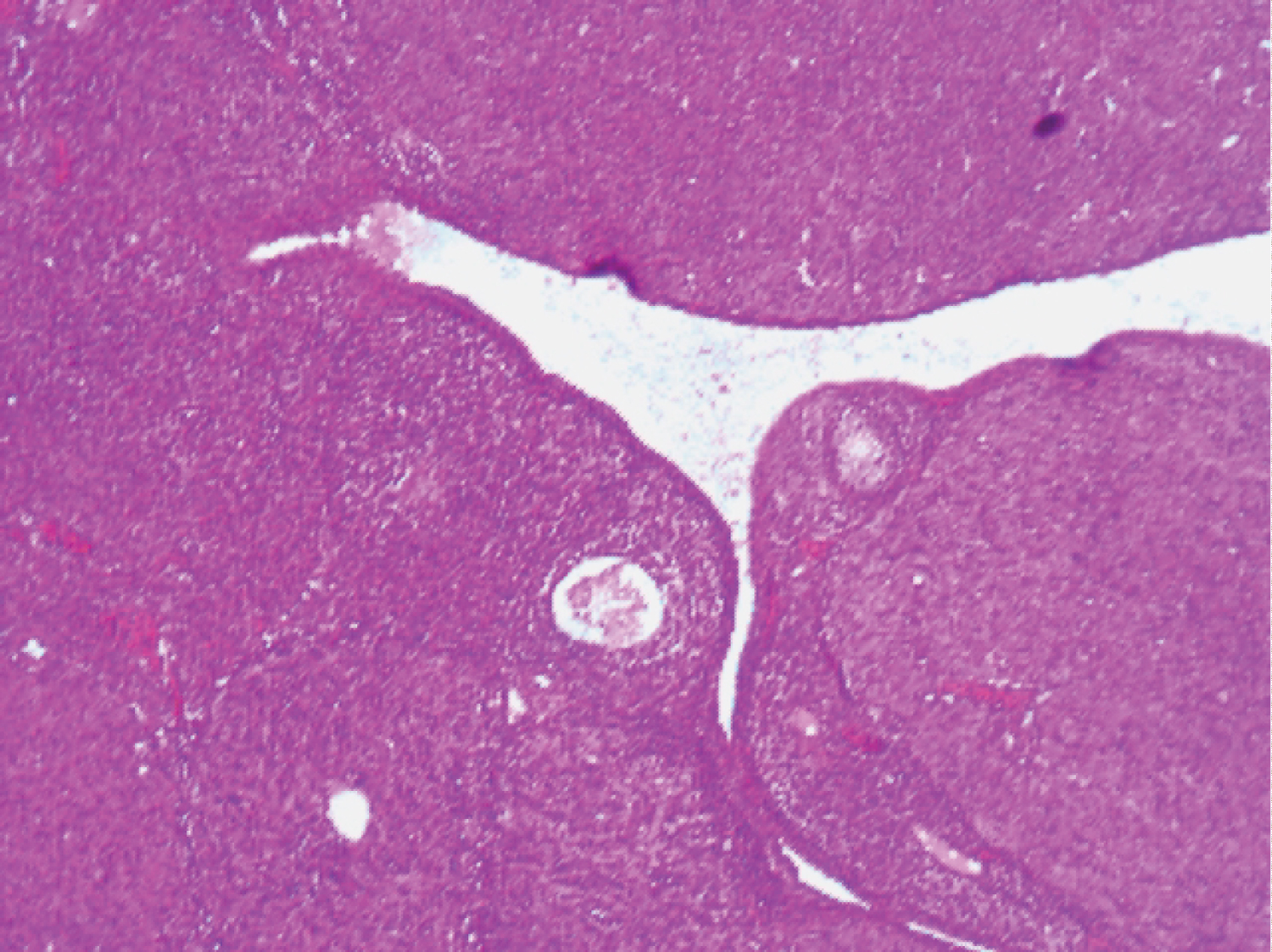 Plate 4: Photomicrograph of section of ovary from Cd plus Fe exposed rat (H and E x100).
View Plate 4
Plate 4: Photomicrograph of section of ovary from Cd plus Fe exposed rat (H and E x100).
View Plate 4
Various studies on the toxic effects of cadmium using different animal models have been carried out (Liu et al. 1995; Gur et al. 1995; Horiguchi et al. 1996; Sidhu et al. 1997; Asagba and Obi, 2005; Asagba, 2009; Nawal et al., 2015) [11,25-30]. This study evaluated the toxic effects of cadmium and iron to elucidate their individual and combined effects on gonads when the animals are exposed via tainted diets.
However, increase in CAT activity observed in ovary of rats exposed concurrently to Cd plus Fe (Table 3) with the attendant decrease in CAT activity in serum of rats exposed to both metals concurrently (Table 2) may be attributed to the induction of stress by Cd in the presence of Fe. Cadmium induced oxidative stress in ovary of rats by increasing the CAT activity, SOD activity and MDA level was reported [31,32]. Decrease in MDA level of ovary of rats exposed to iron-tainted diet suggests that Fe only does not have the capacity to induce oxidative stress in the ovary of rats.
The mechanisms of cadmium toxicity are not clearly understood, but it is believed to act intracellularly, mainly via free radical-induced damage, particularly to the lungs, kidneys, heart, bone, central nervous system and reproductive organs [33]. It has also been reported that Cd may induce oxidative damage in a variety of tissues enhancing peroxidation of membrane lipids due to inhibition of antioxidant enzyme. Others have noted that Cd exposure might cause an increase in antioxidant enzyme activities, leading to lipid peroxidation. The alteration of antioxidant enzyme activities may depend on several factors such as, Cd dose, Cd exposure times, type of Cd administration [34] and the presence of other metals.
The level of cholesterol in serum was also used as an index for stress [35,36,11] and as a possible precursor for steroid hormone biosynthesis in this study. Steroid hormones are biosynthesized from cholesterol. Since reproductive hormones are synthesized from cholesterol, it is a good indicator to evaluate any changes in the steroid hormone identified in this study. The observed decrease in ovary cholesterol levels (Table 1) in rats exposed to Cd, Fe and Cd+Fe tainted diet suggests that cholesterol is likely channelled towards steroid hormones biosynthesis as evidenced by the increase level of estradiol and testosterone in sera of rats (Figure 3 and Figure 4). The decrease in ovary cholesterol levels of rats exposed to the metals singly and combined somehow suggests that, concurrent exposure to cadmium and iron may induce stress in the ovary of rats. This also suggests that Cd can exhibit its gonadotoxic effect in presence of Fe.
Results obtained from this study also show increase in serum level of LH and FSH in the rats after exposure to cadmium and iron for 4 weeks via tainted diet. The increase level of FSH could be as a result of decrease level of cholesterol in the serum or accelerated rate of cholesterol degradation. Variation in the LH level observed in the present study appears to be as a result of the modification activity of these metals in the pituitary gland. Cd has been considered as an important endocrine disruptor in the environment [37,30], its activity may result in variations of plasma LH levels, therefore indicating that the metal may act at the hypothalamic level, modifying the activity of the endogenous clock, changing the mean concentration of LH secreted daily by the pituitary gland [38,30].
Increased estradiol and testosterone levels were observed in rats exposed to Cd (midly) as a separate entity and Cd plus Fe combined treated rats. The mechanism for Cd-induced increase in human testosterone is unknown [39]. The increase in testosterone and estradiol levels observed in this study may be the ability of the metal to perturb the levels of these hormones in the serum or their ability to impair gonadotropin level. Hence, the observed increase in the level of estradiol in Cd-treated and Cd plus Fe treated rats may be due to impaired gonadotropin levels reported by Priya, et al. [40]. Furthermore, enzymes required for the biosynthesis of steroid hormones by the ovary have been shown to be affected by cadmium toxicity [41]. Therefore, increase in the levels of ovarian hormones might be mediated through impaired steroidogenic enzymes or gonadotropin levels [39].
Histological examination of the ovary of experimental rats (Plate 2 and Plate 4) revealed discohesive follicular architecture in the ovary of rats. Ovaries obtained from rats exposed to Cd- tainted diet and Cd plus Fe tainted diet, showed gross follicle disintegration and breakage. The result obtained from this study lends credence to data on cholesterol level in the ovary of rat.
Cd treatment-induced disruption of ovarian histoarchitecture was reported in earlier study of Massanyi, et al. [6]. Previous report of Gurel, et al. [42] revealed ovarian follicular cell damage in Cd-treated female rats. This present study revealed that Cd exposed via tainted diet caused histoarchitectural changes in follicular cells and oocytes, with decrease in number of primary follicles, increase number of atretic follicles and unrecognized oocytes. This observation was in consistent with Massanyi, et al. [43]. Thus, the present study revealed the gonadotoxicity of Cd and its possible consequences in presence of Fe.
These findings suggest that Cd is responsible for the biochemical changes induced in the ovary of exposed rats. Cd-induced stress is apparently raised by the presence of iron. Cd toxicity appears not to be influenced by the presence of iron as shown in serum LH level whereas it does in serum FSH level of rat exposed. If Cd only was able to cause the observed changes in the serum levels of the reproductive hormones analysed, it appears that its toxic effect may be influenced in the presence of iron either by down-regulating or up-regulating the activities of these hormones. Findings from histological examination of the ovaries that there was profound disintegration with follicular damage appear to corroborate the biochemical observations made in this study. In conclusion, cadmium toxicity is apparently enhanced in the presence of iron as evidence by its effects on cadmium-associated alteration in the serum levels of estradiol and testosterone. However, there is need to clarify the interaction of Cd with other metals occurring together in nature.
The authors are grateful to Mr Ben (Pharmacology Department) and Mr Akpeh Kingsley (Biochemistry Department) from the University of Benin for their assistance.