Natural Killer (NK) cell has the capability to immunosensitize and cell-mediated lysis of many types of cancer cells, however, most of the cancer cells are evading the NK cell-mediated lysis by down-regulating the NK cell specific ligands. In the present study, we have evaluated the comparative and synergic effect of spironolactone (SPIR) and avemar (AVE) in enhancing the susceptibility of osteosarcoma cells (MG-63) to NK cell-mediated lysis in vitro. The NK cells used in this study was characterised by the expression of CD56 and NKG2D using flow cytometry. At an optimized concentration, SPIR (60 μM) and AVE (5 mg/ml) treatment showed significant upregulation of the NK cell-specific NKG2D ligands (MICA, MICB, ULP1, and ULP2) on MG-63 cells compared to those in untreated group. To evaluate the NK cell sensitisation towards osteosarcoma cells, MG-63 cells were treated with and without SPIR and AVE for 72 hrs, followed by co-culturing with NK cells [different target and effector ratio (E:T), 1:1, 1:2, 1:5, 1:10 and 1:20] for different time periods (4 and 24 hrs). Cytotoxicity analysis reveals that AVE and AVE+SPIR combination treated MG-63 cells are more susceptible to NK cell-mediated lysis than those SPIR treated in time and dose (E: T ratio) dependent manner. In conclusion, SPIR and AVE are potentially enhancing the NK cell mediate lysis of osteosarcoma cells by unregulated the NK cell specific ligands.
NK cell therapy, AVE, Spironolactone, Immunotherapy, Immunosensitization, NKG2D
Treating osteosarcoma (OS) remains as a challenge attributed to the overall survival rate of 5 years to be 67% and not been improved for the past two decades with existing treatments [1]. Despite, conventional cancer therapies confer survival benefits for OS patients, but they often associated with many side effects and tumor relapse which leads a compromised quality of life. Therefore, there is a desperate need to establish new improved therapeutic strategies to have better treatment outcomes in OS patients. Adoptive immunotherapy is rapidly growing as an alternative to conventional treatment for OS. The effect of anti-tumor immune response is varied as based on the type of adoptive immune cells used. Among them, nature Killer (NK) cells have gained immense attention in immunotherapy for many cancer types, including OS [1-3].
NK cell acts as an antigen-nonspecific lymphocyte which plays an important role in recognizing and eliminating virally infected, stressed and malignant cells [4]. NK cells, among the various immune cells, exhibit the highest cytotoxic capacity and have an ability to induce lysis of cancer cells without the need for any prior sensitization [5]. The cytotoxic effects are determined by the proportional ratio between activated receptors of NK cells and their respective ligands on target cells. NKG2D, one of the main activating receptors in NK cells, recognises wide range of ligands including MICA/B and ULBPs which are highly expressed in several cancer cells [6-9]. However, most of the cancer cells evades NK cell-mediated lysis by down-regulating the NK cell-specific ligands, therefore resulted in resistant to NK cells mediated lysis. Thus, needed strategy or therapeutic molecules that can regulate (activate or induce) the NKG2D ligands on the target cells to enhance the therapeutic effect of NK cell-mediated lysis would be relevant and necessitate.
Spironolactone (SPIR) is an antimineralocorticoid drug, and most commonly used to treat heart failure, liver scarring and kidney disease [10]. Recently, Leung, et al. reported that SPIR induced expression of NKG2D ligands in different solid tumors through activation of ATM-Chk2 mediated checkpoint pathway, suggesting SPIR most likely to induce susceptibility of cancer cells to NK cell-mediated lysis [11]. Similarly, AVE is a product of fermented wheat germ extract and commonly used as a natural, non-toxic nutritional supplement for cancer patients to improve their quality of life. It contains two quinones; as 2,6-dimethoxy-1,4-benzoquinone (DMBQ) and 2-methoxy-benzoquinone that likely to take part in several biological activities such as antiproliferative, antimetastatic and immunological effects [12-14]. Although AVE and SPIR have been reported to induce a wide range of biological effects through various mechanisms, their immunomodulatory effect through modulation of NKG2D ligands expression in OS remains to be explored.
To this aim, here we have demonstrated that SPIR and AVE individually and synergistically up-regulate the expression of NKG2D ligands in OS cell line. Our results demonstrated that the synergetic effect of SPIR and AVE enhanced the susceptibility of OS cells to NK cell-mediated lysis by up-regulating the NKG2D ligands. Overall, our data suggest that the combination of AVE plus SPIR could be an effective approach to enhance therapeutic effect of NK cell-mediated lysis in OS.
After informed consent was obtained from all three (3) donors, 30 ml of peripheral blood (PB) was collected in accordance with local and national regulatory guidelines. The procedures followed were in accordance with the ethical standards described by the Helsinki Declaration (64th WMA General Assembly, Fortaleza, Brazil, October 2013). The peripheral blood mononuclear cells (PBMCs) which contain NK cells were isolated by lymphoprep (AXIS-SHIELD PoC AS, Oslo, Norway) density gradient medium and centrifugation at 2000 rpm for 20 mins. Isolated PBMCs were washed twice with phosphate buffered saline (PBS) and the cell pellet was resuspended in culture media containing 10% autologous plasma and IL2. Suspended PBMCs were cultured as per the protocol described earlier [15] and cells were cultured for the period of 14 days.
The expanded NK cell phenotype and purity were assessed by employing markers via CD56 and NKG2D. Immunostaining was performed as per manufacturer's instruction. Stained cells were re-suspended in PBS and analyzed by using flow cytometry (FC-500 Beckman Coulter, USA). The acquired data were analyzed using CXP software provided by the manufacturer.
The Osteosarcoma cell line (MG-63) was provided by University Technology MARA, Shah Alam, Malaysia. Cells were cultured according to the recommendation of the supplier, in DMEM containing 10% FBS (Invitrogen, BRL, Burlington, ON, Canada). Cells were cultured without any stimulatory supplements in a humidified incubator at 37 ℃ with 5% CO2 . Upon 70-80% confluency, the adherent cells were passaged by trypsinization (0.25% trypsin-EDTA, Invitrogen, BRL, Burlington, ON, Canada) and consumed for downstream experiments.
The MG-63 cells were seeded in 96-well plates at a density of 3 × 103 cells/well and incubated for 24 hrs. Thereafter, the medium was replaced, and incubation continued for 72 hrs in the presence or absence of the drugs (Spironolactone and AVE) of the various concentrations. Upon 72 hrs MTT (3-(4, 5-dimethylthiazol- 2, 5-diphenyl tetrazolium bromide) (Sigma Chemical Co., St. Louis, MO, USA) proliferation assay was carried out to assess drug toxicity. MTT was dissolved in PBS at 5 mg/ml; added to the culture medium at a dilution 1:10 prior to incubation at 37 ℃ for 4 h. The medium was aspirated; 100 μl of DMSO was added/well and the 96-well plate was read using a microplate reader (DYNEX Technologies, Chantilly, VA, USA) at the wavelength of 570 nm.
The MG-63 cells were seeded at density of 6 × 103 cells/cm2 and incubated for 24 h, the medium was replaced by a culture medium that contained SPIR (60 μM/ml) (Sigma Chemical Co., St. Louis, MO, USA) and AVE (5 g/ml) and the cells were incubated for 72 h. The morphological changes were assessed using an inverted microscope.
Total RNA was extracted from MG-63 cells with and without treatment of SPIR (60 μM/ml) and AVE (5 mg/ml) by using RNeasy Mini Kit (QIAGEN). The preparation of the first-strand cDNA was conducted following the instruction of the QuantiTect (Qiagen). RT-PCR was performed using the QuantiTect Rev.Transcription Kit (QIAGEN, Valencia, CA, USA). Genes of interest were obtained using primers synthesized from AIT biotech (Singapore) as shown in Table 1. Gene expression was quantified by ImageJ software and the gene expression was normalized with housekeeping gene, GAPDH.
Table 1: List of primers used for RT-PCR. View Table 1
The MG-63 cells (Target cell) were seeded in 96-well plates at a density of 3 × 103 cells/well and incubated for 24 h. Thereafter, the medium was replaced, and incubation continued for 72 h in the presence or absence of drugs [SPIR (60 μM/ml) alone, AVE (5 g/ml) alone and SPIR (60 μM/ml) along with AVE (5 g/ml)]. The media was removed and washed with PBS after 72 hrs, then NK cells (Effector cells) were seeded at different ratio (T:E), 1:1, 1:2, 1:5, 1:10 and 1:20] for different time periods (4 hrs and 24 hrs). MTT test was then performed after the plates were centrifuged 1500 RPM for 5 mins. Cytotoxicity was calculated using the formula [1-(Mean OD of co-cultured cells-Mean OD of Effector cells)/Mean OD of control cells] × 100%. Triplicate experiments were performed and average values with mean ± SEM were represented.
All data were statistically analyzed using Student t-test. Data were presented as the mean ± standard deviation for cytotoxicity and RT-PCR assay. P < 0.05 was considered to be statistically significant.
As revealed earlier NKG2D-NKG2DL pathway plays a crucial role in NK cell-based therapy to eliminate the cancer cells. In order to that, we first evaluate the purity and anti-tumor functionality of NK cells using unique biomarkers CD56 and NKG2D respectively, assessed by flow cytometry. The analysis revealed that purity and anti-tumor functionality of ex-vivo expanded NK cells were more than 95% (Figure 1). These results confirmed that NK cells were successfully activated and expanded without altering their original characteristic of anti-tumor effect.
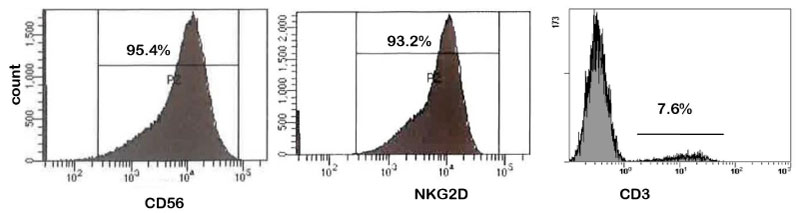 Figure 1: Flow cytometry analysis of human peripheral blood derived NK cells.
Figure 1: Flow cytometry analysis of human peripheral blood derived NK cells.
Ex-vivo expanded NK cells were expressed more than 95% of CD56 and NKG2D which directly correlated with their purity and anti-tumor potency, respectively.
View Figure 1
Cell growth inhibitions were assessed with a range of concentration starting from 20 μM to 200 μM of SPIR and 200 μg to 2000 μg of AVE. It was noted that the median effective dose (ED50) was 100 μM/ml and 5 mg/ml of SPIR and AVE respectively (Figure 2a and Figure 2b). MG-63 cells were cultured and treated with SPIR (60 μM/ml), AVE (5 g/ml) SPIR (60 μM/ml) plus with AVE (5 g/ml) and the cells were incubated for 72 hrs. After 72 hrs of incubation, there was no morphological changes noted in cells treated with SPIR (60 μM/ml) (5 g/ml) and SPIR (60 μM/ml) plus with AVE (5 g/m (Figure 2b, Figure 2c and Figure 2d).
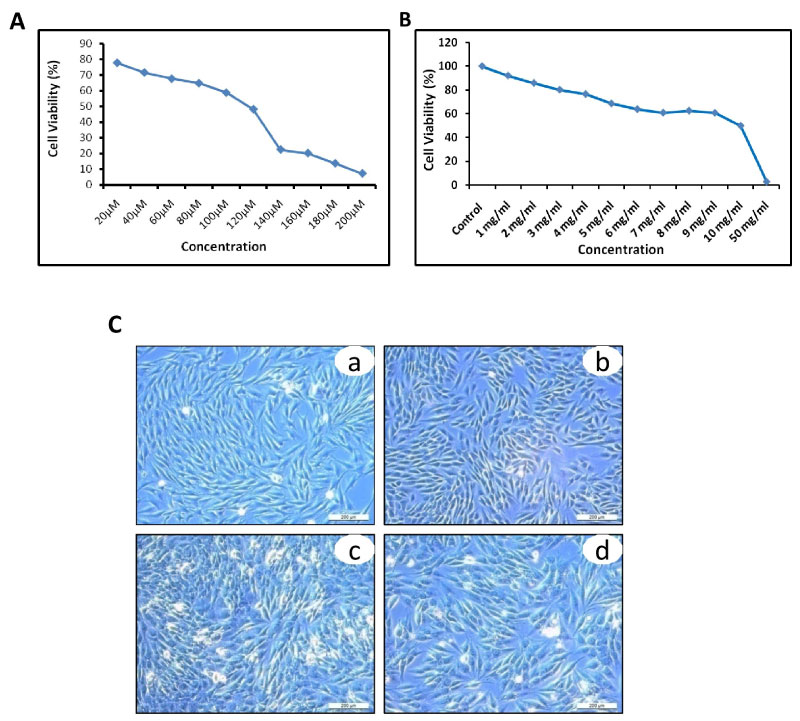 Figure 2: Cytotoxicity of SPIR and AVE on osteosarcoma cell line (MG-63).
Figure 2: Cytotoxicity of SPIR and AVE on osteosarcoma cell line (MG-63).
SPIR and AVE effectively inhibit the OS cell proliferation at dose and time dependent manner: a, b) The median effective dose of SPIR and AVE was founded at the concentration 100 μM and 5 mg/ml, respectively; c) SPIR and/or AVE could not alter any significant morphological changes on OS cells: (a) Untreated; (b) SPIR treated; (c) AVE treated and (d) SPIR and AVE treated OS cells.
View Figure 2
An optimized concentration of SPIR (60 μM) and AVE (5 mg/ml) were used to treat MG-63 cells in culture and mRNA expression of NKG2D ligands (MICA, MICB, ULP1 and ULP2) were compared with MG-63 control group. The expression level of MICA, MICB, ULP1, and ULP2 significantly higher in cells treated with SPIR, AVE, and SPIR+AVE compare with untreated group (Figure 3a and Figure 3b). On the other hand, the expression of MICA/B (3.3 ± 0.8, 2.4 ± 0.4 vs. 3.1 ± 0.2, 1.7 ± 0.2 vs. 2.9 ± 0.1, 1.8 ± 0.7) was positively expressed at higher levels in cells treated with AVE alone compared with cells treated with SPIR alone and SPIR+AVA. Whereas, mRNA expression of ULBP2 in cells treated with AVE alone was expressed equally to cells treated with SPIR alone and SPIR+AVA (7.8 ± 0.6 vs. 7.9 ± 0.0 vs. 7.7 ± 0.5) and significantly dropped the expression level of ULBP1(1.4 ± 0.3 vs. 1.5 ± 0.5 vs. 1.8 ± 0.4). The results demonstrated that the cells treated with SPIR and AVE show higher expression of NKG2D ligands compare with control. However, the overall results demonstrated that the cells treated with AVE alone show better expression level compared with control and cells treated with SPIR alone and SPIR+AVE (Figure 3b).
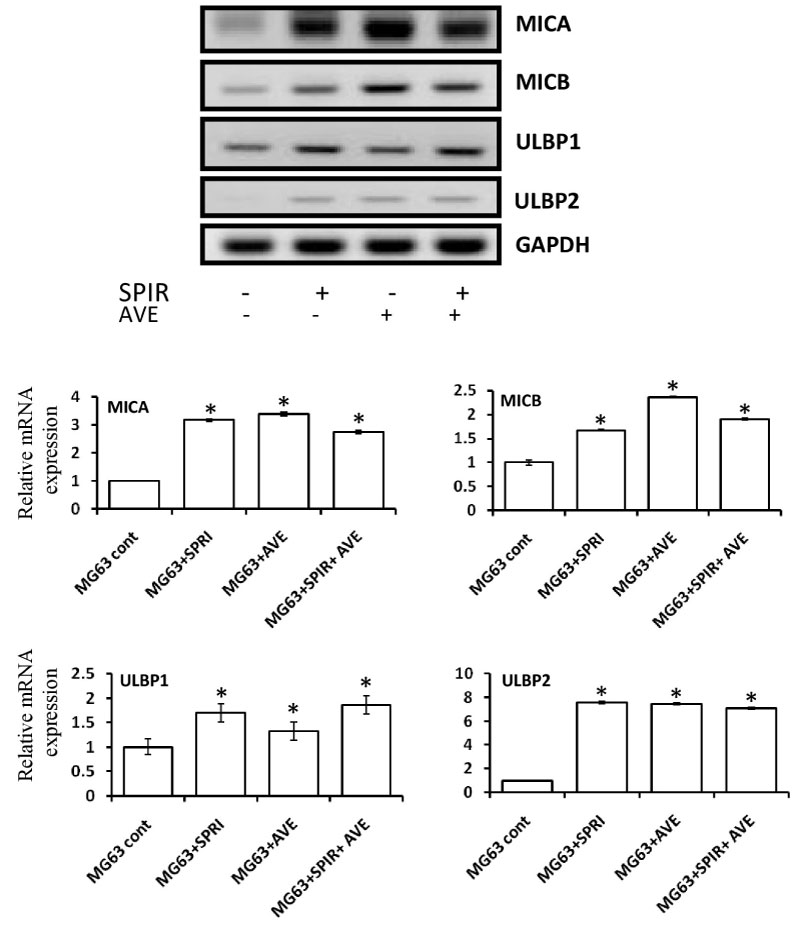 Figure 3: RT-PCR analysis of NKG2D ligands expression in SPIR and AVE treated and untreated osteosarcoma cell line (MG-63).
Figure 3: RT-PCR analysis of NKG2D ligands expression in SPIR and AVE treated and untreated osteosarcoma cell line (MG-63).
SPIR and AVE Potentially up-regulated the NKG2D Ligands on targeted OS cell line. Electrophoresis gel images and their respective densitometry clearly revealed that SPIR and AVE significantly up-regulate the NKG2D ligands on OS cells. The variation within each set of triplicates is shown with mean of SD ± *P < 0.05 (n = 3).
View Figure 3
We have carried out triplicate experiments of dose-dependent and time-dependent. First, we have categorized the target into four groups MG-63 alone MG-63 treated with SPIR MG-63 treated with AVE and MG-63 treated with SPIR+AVE. We have seeded 3 × 103. After 24 of cell seeding, the condition of attached cells onto the surface of multi-well was observed when the media was replaced with culture media alone, culture media with SPIR, AVE, and SPIR+AVE respectively. After 72 h incubation, Effector cells were seeded at 1:1, 2:1, 10:1, and 20:1 ratios. We set to evaluate OD measure of the cell cytotoxicity after 4 hrs and 24 hrs of coculture. The cytotoxic activity was then evaluated with MTT-assay. Hence, the assessment of cytolytic activities of NK cell lysis against MG-63 cancer cell at different- time points and target ratios were measured (Figure 4a and Figure 4b). After four hours of coculture, NK cells completely killed all target cells in cells treated with SPIR+AVE (1:1 to 20:1) and the cytolysis activity was 100 ± 0.1%.Whereas, 100 ± 0. cytolytic activity was noted only at 20:1 in cells treated with SPIR, AVE and without treatment. However, the cytolytic activity was better in cells treated with AVE alone compare with cells treated with SPIR alone and without treatment. The cell lysis was > 88 ± 0.9% were the cells co-cultured from at 1:1, 2:1, 5:1 10:1 and 20:1 ratios. On the other hand, they cytolysis activity was increased when the cells were maintained for 24 hrs. The target cells were completely killed at 5:1 ratio in cells treated with SPIR, AVE and SPIR+AVE, whereas 90 ± 1.8% cells only targeted at 5:1 ratio in cells without treatment (Figure 2b). The overall results confirmed that the NK cell cytolysis activity was happened dose and time depended manner. The ratio 5:1 will be the ideal dose cells with or without treatment to eradicate the target cells.
 Figure 4: Cellular cytotoxicity of NK cell on SPIR and AVE treated and untreated osteosarcoma cell line. SPIR and AVE treatment increased the susceptibility of OS cells to NK cell mediated lysis: a) 4 hrs cytotoxicity of NK cells vs. SPIR and/or AVE pre-treated OS cells; b) 24 hrs cytotoxicity of NK cells vs. SPIR and/or AVE pre-treated OS cells. The variation within each set of triplicates is shown with mean of SD ± *P < 0.05 (n = 3).
View Figure 4
Figure 4: Cellular cytotoxicity of NK cell on SPIR and AVE treated and untreated osteosarcoma cell line. SPIR and AVE treatment increased the susceptibility of OS cells to NK cell mediated lysis: a) 4 hrs cytotoxicity of NK cells vs. SPIR and/or AVE pre-treated OS cells; b) 24 hrs cytotoxicity of NK cells vs. SPIR and/or AVE pre-treated OS cells. The variation within each set of triplicates is shown with mean of SD ± *P < 0.05 (n = 3).
View Figure 4
Survival of OS patients has not improved in 20 years, furthermore, the prognosis of the patient is very poor; it's due to the high frequency of diagnosis with the metastatic or late stage of disease and limited therapeutic effects on current treatments. Therefore, novel therapeutic strategies are greatly required for treating OS [16,17]. Immunotherapy, especially NK cell-based therapy has gained special attention to treat many cancer types, including OS. In agreement with a previous study reports that circulatory NK cells were significantly dropped in patients with OS than a normal individual, suggesting that NK cell play a vital role in prevention and elimination of OS cells [18]. The anti-tumor immune response of NK cell has usually been determined by the interaction between the NKG2D receptor on NK cells and NKG2D ligands on target cells. By shedding or down-regulating the NKG2D ligands, cancer cells are evading NK cell-mediated lysis [19,20]. Thus, it is indeed to identify the methods to induce or up-regulate NKG2D ligands on the target cell to enhance the NK cell cytotoxicity.
In line of that, we have selectively chosen two immunomodulatory drugs SPIR and AVE which were recently reported to exert anti-tumour response in various tumours types by up-regulating the NK specific ligands and/or suppressing the invading mask of MHC-1 on target cells and resulting in increased immunotherapeutic effect of NK cells [3,11,13,21]. Our cytotoxicity assay revealed that SPIR and AVE did not inhibit the proliferation of OS cells at the concentration of 60 μM and 5 mg/ml, respectively. Furthermore, there were no significant morphological changes noted on treated (at the specified concentrations of SPIR and AVE) cell line. Next, we evaluated effect of SPIR and AVE in up-regulating the NKG2D ligands to exhibit anit-tumour response of NK cells on OS cells. MICA, MICB, ULBP1, and ULBP2 are primary NKG2D ligands which can potentially activate the NKG2D receptor on NK cells and result in enhancing the NK cell-mediated tumor clearance [22]. Therefore, we analyzed the impact of SPIR and/or AVE on the expression of MICA/B and ULBP1/2 in OS cells. Our RT-PCR analysis revealed that the mRNA expression of MICA/B and ULBP1/2 were significantly up-regulated by SPIR and/or AVE (Figure 3). Especially, MICA and MICB were certainly up-regulated by AVE alone than those of cells treated with SPIR alone or SPIR+AVE which reflect that AVE treated cells are more susceptible to NK cell-mediated lysis. There was a quite few number of cell signaling pathways identified to regulate the MICA and MICB in different cancer cells, for example HER2-HER3 signaling in breast cancer [23], Erk signaling in myeloma and ovarian cancer [24,25] and ATM-Chk2 pathway in colon cancer [11]. On another hand, ULBP1 and ULBP2 were remarkably up-regulated by SPIR alone or SPIR+AVE as compared to AVE alone. These data suggested that in comparison to AVE, SPIR tightly regulate the ULBP1 and ULBP2 expression rather than MICA and MICB and these observations are in agreement with one previous report [11] but they haven't compared with each other.
Conversely, NK cell cytotoxicity was determined not only by the expression pattern of NKG2D ligands on the target cell, it also the expression pattern of NKG2D receptor on NK cells [1]. NKG2D act as a primary activation receptor in most of the activated NK cells and engagement of NKG2D alone is more sufficient to enhance the NK cell-mediated lysis. Thus, production of activated NK is desperately needed to achieve the good clinical outcome. We have validated our standard protocol for NK cell production through flow cytometry analysis using respective markers. Our data shows that the surface expression of NKG2D and CD56 on NK cell was greater than 95% and these results demonstrated that ex-vivo activated and expanded NK cell was successfully manufactured at high purity along with an anti-tumour response [15].
A new generation of cancer drugs which directly or indirectly induce the NKG2D ligands on targeted cancer cells is an effective therapeutic strategy for treating cancer with resistance nature [26-28]. Although our finding showed that SPIR and/or AVE potentially up-regulated the NKG2D ligands on OS cells, but their sensitivity to NKG2D expressed NK cell-mediated lysis is not explored. In order to that, we have investigated the effect of SPIR and AVE on NK cell-mediated lysis of OS cells. Our cellular cytotoxicity analysis showed that SPIR and/or AVE effectively enhanced the OS cells susceptibility to ex-vivo activated and expanded NK cells in dose (E/T ratio) and time-dependent manner, which may rely on an NKG2D-NKG2D ligands dependent interaction between NK cells and OS cells. Recently, Barisone, et al. proven that AVEMER contain benzoquinones protein which enhance for NK cell activation [15]. Furthermore, Guma, et al. [1] also reported that NK cell-mediated lysis was dramatically dropped by blocking either NKG2D ligands or NKG2D receptor. These observations apparently demonstrated that NK cells are most aggressively eliminate the cancer cell through NKG2D-NKG2DL interaction.
In summary, overall our results demonstrated that SPIR and AVE individually or synergistically enhance the immune sensitization and susceptibility of OS cells to NK cell-mediated lysis by up-regulating NKG2D ligands on OS cells. Moreover, we provide the evidence to confirm that AVE alone or combination with SPIR enhance the NK cell-mediated lysis of OS cells. Altogether, our data suggest that AVE and SPIR could be used as immunomodulatory drugs to enhance the therapeutic effect of NK cells in the patient with OSteosarcoma [29].
The authors express no conflicts of interest towards the publication of this paper.