One of the main components of Pomegranate Seed Oil is punicic acid, an omega-5 fatty acid that is capable of inhibiting breast and prostate cancer cell proliferation, although the mechanisms of action are not yet known. In order to further understand its potential anti-cancer properties, we investigated the effect of punicic acid on proliferation and cytotoxicity of human MCF-7 breast cancer cells. Cells were treated with punicic acid for up to 72 hours, and cell proliferation, cytotoxicity, and apoptosis were measured. We found that 10 and 50 ug/ml punicic acid significantly reduced viable cell density by 72 hours as measured by MTS assay. Similarly, we found that both concentrations significantly increased cytotoxicity by this time point, as measured by an LDH assay. Cell death was confirmed to be, at least in part, due to apoptosis, as detected by Annexin V and propidium iodide fluorescence. Since many of the peroxiredoxin (Prdx) family of antioxidant proteins are known to be involved in cancer cell survival, and our lab previously demonstrated their upregulation in MCF-7 cells, we also examined the effect of punicic acid on the Prdx mRNA expression. We found elevated levels of Prdx1-4 in punicic acid treated cells, with statistically significant induction of Prdx1 and Prdx4. Altogether, our data definitively demonstrates anti-cancer properties of punicic acid, as well as its ability to regulate antioxidant gene expression in these cells. This research has important implications in the field of natural products as therapeutic options for cancer treatment, as opposed to conventional chemotherapy.
Breast cancer is one of the most common cancers in women worldwide, and the leading cause of cancer deaths in women. Over 1.5 million women are diagnosed with breast cancer every year throughout the world [1]. Breast cancer metastasis to the bone, lungs, liver and brain often accounts for the incurability of the disease. In the United States, 80% of breast cancer patients are expected to live five years after their initial diagnosis if the tumor is detected early [2]. Risk factors that contribute to the onset of disease include aging, family history, estrogen, lifestyle and sex. Most breast cancers occur in women, with the number of cases being 100 times higher than in men [3].
A common feature of cancer cells is elevated levels of Reactive Oxygen Species (ROS), a large group of highly reactive oxygen-derived small molecules that include radical and non-radical species [4,5]. ROS can react with lipids, proteins and nucleic acids which elicit tissue damage leading to necrosis and pro-apoptotic pathways. In many cancers, oxidative stress can occur due to the imbalance between the excess production of free radicals and the cellular antioxidant defense [6,7]. Compared to normal epithelial cells, breast cancer cells have higher levels of ROS and oxidative stress, which can trigger genomic instability and gene dysregulation. In fact, higher ROS has been shown to promote carcinogenesis and lead to an increased likelihood of cell proliferation, metastasis, angiogenesis and proliferation in breast cancer cell lines [8].
Antioxidants that protect cancer cells from ROS-induced damage can be endogenous (made naturally by the cells and tissues) or exogenous (retrieved from the environment). Both endogenous and exogenous antioxidants can reduce the harmful effects of ROS, ultimately enhancing the immune defense and lower the risk of disease and cancer [8,9]. Exogenous antioxidants from fruits and vegetables are of major biomedical interest due to their well-documented health benefits. The pomegranate (Punica granatum L.) fruit, from the family Punicaceae, has been shown to have high antioxidant capacity in human and animal models [10]. Pomegranates can prevent various inflammatory-mediated diseases through various biological actions. Pomegranate Seed Oil (PSO) specifically, has been shown to possess anti-carcinogenic properties in an array of different cancer types including skin, breast, and prostate and colon [10-15]. PSO comprises 12%-20% of the total seed weight, and primarily consists of triacylglycerols. The hydrophilic components of PSO are known to possess a broad spectrum of biological activities, which include regulating gene expression and suppressing chemically induced carcinogenesis [16].
Compounds extracted from the Pomegranate seed contain conjugated octadecatrienoic fatty acids. One such fatty acid, punicic acid (PA), is an omega-5 long chain polyunsaturated fatty acid that makes up approximately 65-80% of the oil from pomegranate seeds [11]. Punicic acid has been found to prevent cancer proliferation and induce cytotoxicity in MCF-7 mammary cancer cells and increase apoptosis of the MDA-MB-435 human breast cancer cells [17,18]. The mechanism of action of punicic acid on MCF-7 breast cancer cells is not well understood, but researchers believe punicic acid enters the cell and is utilized in the synthesis of diacylglycerol (DAG). Combined with lipid peroxidation, diacylglycerol activates protein kinase C (PKC), which participates in signal transduction cascades that leads to inhibition of proliferation and activation of apoptosis [17,18].
To better understand the effect of punicic acid on breast cancer cells and its role in gene dysregulation, we sought to examine its effect on MCF-7 viability, apoptosis, and the expression of the Peroxiredoxin genes, a family of thiol-specific antioxidant genes that are upregulated in many cancers. This work has pertinent implications in the field of natural products as a therapeutic option for cancer treatment.
The human MCF-7 breast cancer cell line was purchased from ATCC. The cells were cultured in EMEM media containing 5% Fetal Bovine Serum (FBS) ATCC 30-2020 and 50 ng/ml insulin. Cells were grown and maintained at 37 ℃ in a humidified 5% CO2 incubator. Punicic Acid was purchased from Cayman Chemicals (Cat# 26057) as a 10 mg/ml stock dissolved in methanol. For all cell experiments, viable cells were quantified by hemacytometer using a trypan blue exclusion assay. Viable cells were seeded into appropriate well plates for each assay and treated with 10 or 50 ug/ml punicic acid (or an equivalent volume of 70% ethanol as a control). The stock punicic acid was stored at -20 ℃ and protected from light.
To measure cell growth and viability, cells were seeded at 0.5 × 105 cells/mL in a 24 well plate in replicates of 4-6. Cells were treated with varying concentrations of punicic acid or ethanol control and incubated for 72 hours. After 72 hours, media was removed and cells were assayed for MTS conversion using the Cell Titer Cell Proliferation assay (Promega).
To measure cytotoxicity, MCF-7 cells were plated at a density of 0.5 × 105 cells/mL in a 24 well plate in replicates. Cells were treated with varying concentrations of punicic acid or ethanol control and incubated for 72 hours. At 24, 48, and 72 hours post-treatment, media was removed and tested for LDH levels using the colorimetric Lactate Dehydrogenase (LDH) Assay (Promega), which quantifies cellular cytotoxicity by measuring the release of LDH into cell culture media when the plasma membrane is damaged.
To measure apoptosis, MCF-7 cells were plated at a density of 0.5 × 105 cells/mL in a 24 well plate in replicates of six. Cells were treated with punicic acid or ethanol control and incubated for 72 hours. Cells were rinsed with 1X PBS and assayed using the Annexin apoptotic assay (ThermoFisher), which includes a green fluorescent labeling of membrane Annexin V, and propidium iodide counterstaining of nuclei. Cells were imaged using a fluorescence microscope.
To measure Peroxiredoxin expression, MCF-7 cells were plated at a density of 0.5 × 105 cells/mL in a 24 well plate in replicates of three. Cells were treated with 10 ug/ml punicic acid or ethanol control for 24 hours and RNA was isolated using the RNAqueous-4PCR Kit (ThermoFisher), and subsequently reverse transcribed with Reverse Transcriptase enzyme and buffer (Applied Biosystems). Taqman Real Time PCR Assays were conducted in triplicate using Applied Biosystems primer/probes specific to each of the six mammalian peroxiredoxin genes. These included human-specific TaqMan assays for Prdx1 (Hs00602020), Prdx2 (Hs03044902), Prdx3 (Hs00428953), Prdx4 (Hs00197384), Prdx5 (Hs00738905) and Prdx6 (Hs00705355). An 18S rRNA primer/probe (Applied Biosystems cat#4333760F) was used as an endogenous control for all RT reactions, and relative quantification (RQ) was calculated.
The means of individual treatment groups in each experiment were statistically compared using a two-tailed student's t-test, assuming equal variances.
In order to determine if punicic acid had an anti-proliferative effect on MCF-7 cells, we treated cells with two different concentrations of punicic acid (10 ug/ml and 50 ug/ml) or ethanol control for 72 hours and measured the remaining viable cells using an MTS assay. As is shown in Figure 1, both concentrations of punicic acid significantly reduced viable cell number after 72 hours, resulting in about a 50% reduction. We saw no statistically significant difference between the effects of the two punicic acid concentrations.
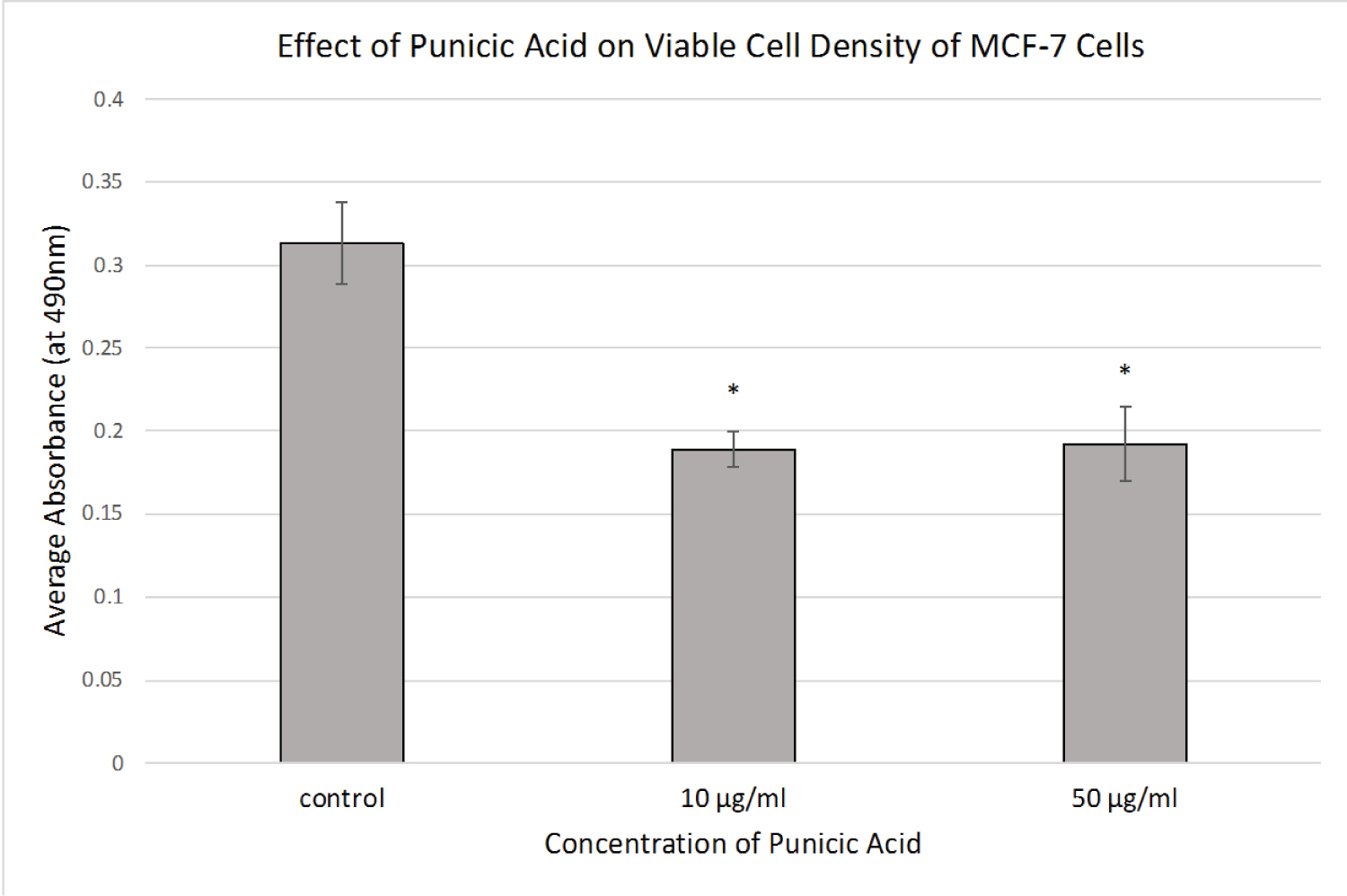 Figure 1: Effect of Punicic Acid on MCF-7 Viable Cell Density.
Figure 1: Effect of Punicic Acid on MCF-7 Viable Cell Density.
MCF-7 cells were treated with ethanol control or punicic acid at the indicated concentrations for 72 hours, and viable cell density was measured using an MTS assay. Average absorbances are shown (+/- stdev). *p < 0.05.
View Figure 1
To further elucidate whether the observed MTS results were due to growth inhibition or cell death (or both) we examined the effect of the treatments on LDH release, which is a measure of cytotoxicity. We measured LDH release at 24, 48 and 72 hours post-treatment. As shown in Figure 2, cells treated for 48 hours with a low concentration of punicic acid had a significant increase in cytotoxicity, while cells treated for 72 hours had an even higher level of cytotoxicity with both concentrations of punicic acid. The modest increase in cytotoxicity in the control cells was shown to be due to overgrowth (as is evident by cell morphology, (Figure 3)).
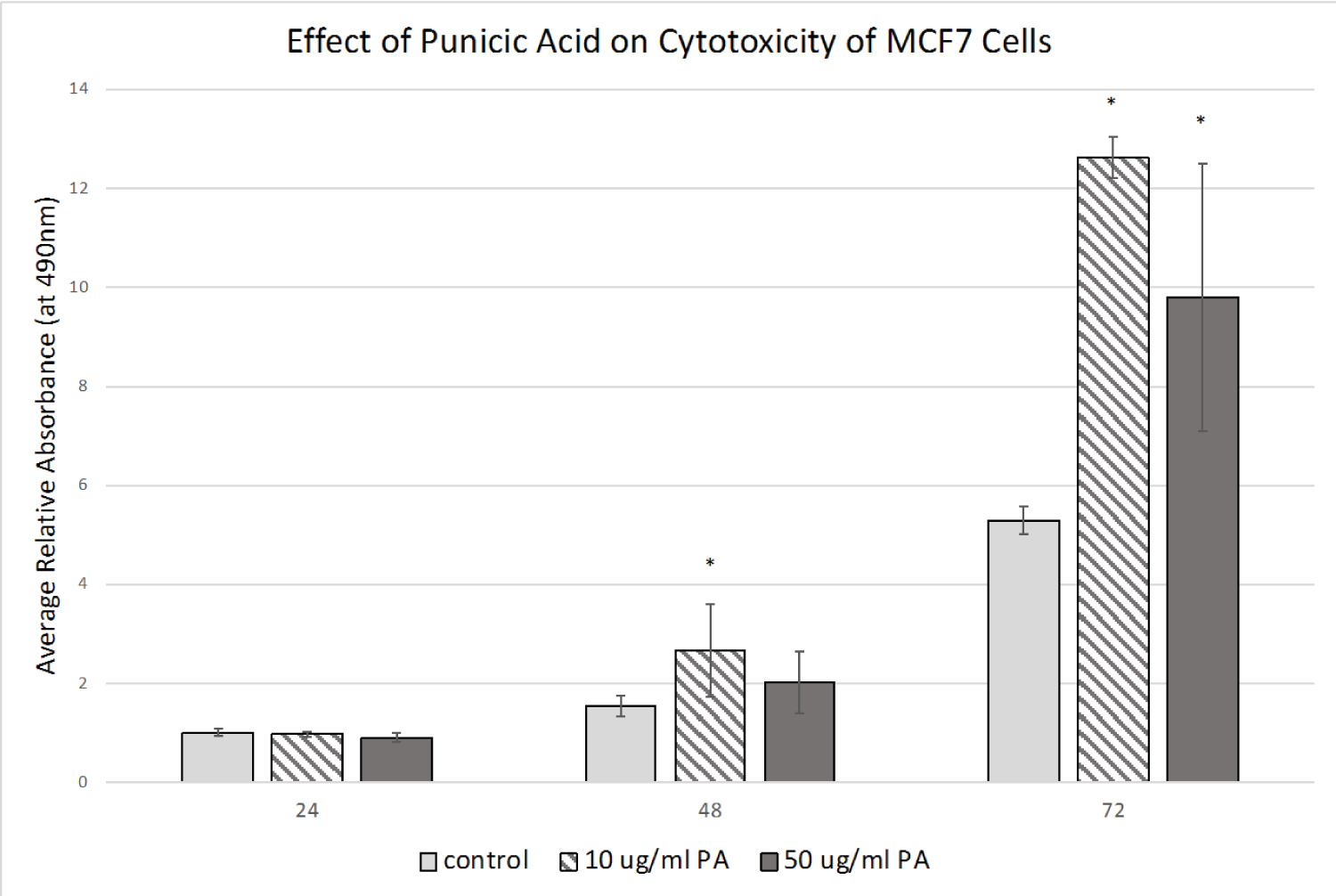 Figure 2: Effect of Punicic Acid on MCF-7 Cytotoxicity.
Figure 2: Effect of Punicic Acid on MCF-7 Cytotoxicity.
MCF-7 cells were treated with ethanol control or the indicated concentrations of punicic acid (PA) for 72 hours and released LDH was measured in the media at 24, 48, and 72 hours post treatment. Average absorbances (relative to 24-hour controls) are shown (+/- stdev). *p < 0.05 (relative to control for each time point).
View Figure 2
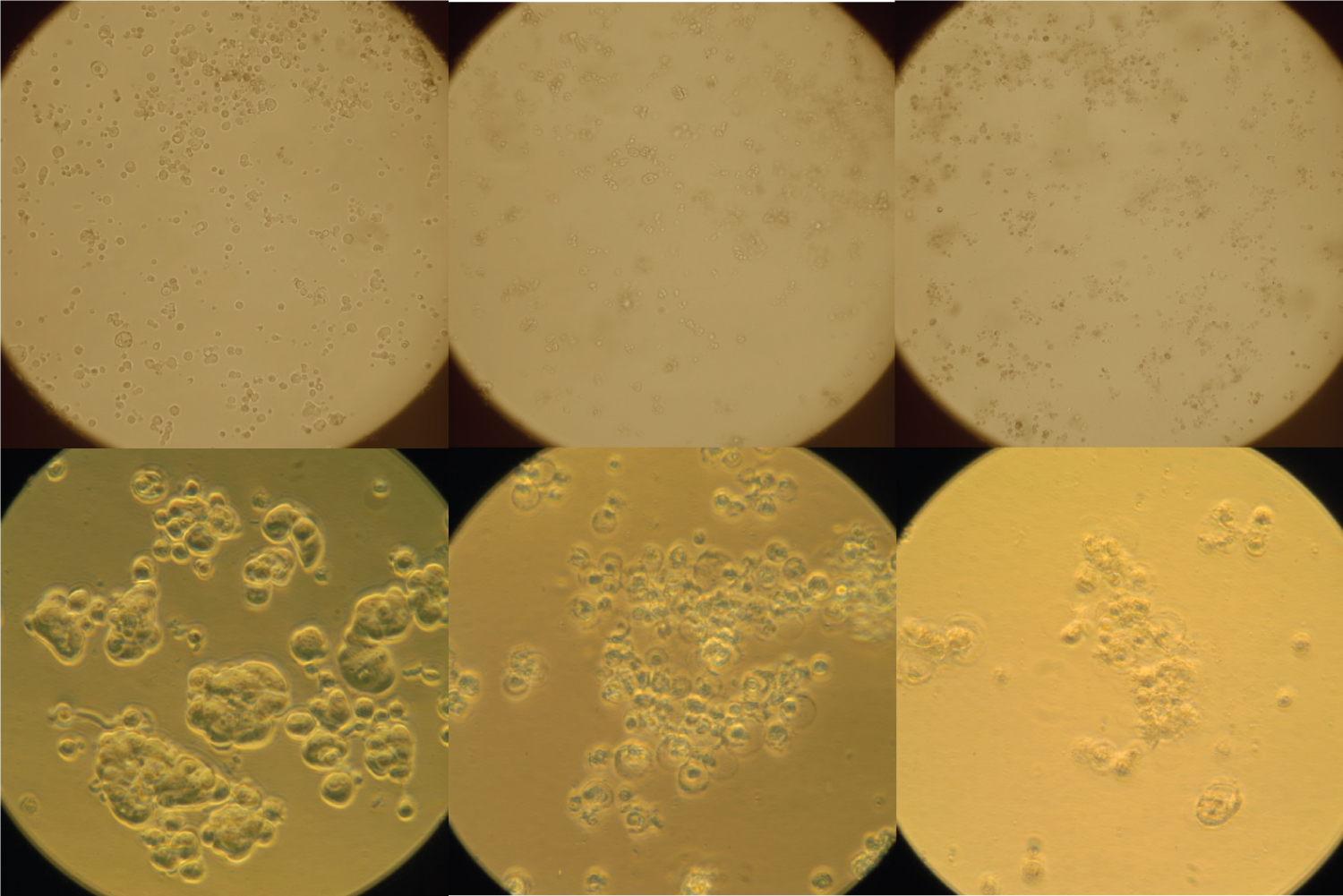 Figure 3: Effect of Punicic Acid on Cell Morphology.
Figure 3: Effect of Punicic Acid on Cell Morphology.
MCF-7 cells were treated with ethanol control (left), 10 ug/ml punicic acid (middle) or 50 ug/ml punicic acid (right) for 72 hours and photographed with phase contrast microscopy at 100 X (top) and 400 x (bottom).
View Figure 3
We examined the morphology of the cells in response to punicic acid to better understand the cell-level effects. In Figure 3, phase contrast images are shown for each treatment type after 72 hours. Microscopy images at 100X show a clear reduction in cell density in both experimental treatment groups. At 400X, morphological changes in the cells treated with punicic acid are apparent, including a reduction in size, increase in granulation, and membrane blebbing and nuclear fragmentation characteristic of apoptosis. Together, these data confirm the cytotoxic effects of punicic acid and suggest a role for apoptosis in this cell death.
We sought to measure apoptosis in punicic acid-treated MCF-7 cells and used the Annexin V/Propidium Iodide assay. Cells were treated as described above and assayed at 48 hours for apoptosis. Cells were imaged for phase contrast, green fluorescence (Annexin V staining) and red fluorescence (nuclear PI staining). That time frame was chosen so cells in earlier stages of apoptosis could be detected, since late-stage apoptosis and cells post-apoptosis may no longer be detected at the 72-hour time period. As shown in Figure 4, 48 hours of treatment with 10 ug/ml punicic acid showed a marked increase in PI-positive (dead) cells. Many of these cells also exhibited membrane-localized Annexin V staining, indicating apoptotic cells. With 50 ug/ml punicic acid treatment, there was a clear reduction in the number of intact cells by 48 hours, as shown by phase contrast microscopy. Virtually all of the remaining cells were positively stained for propidium iodide and membrane-localized Annexin V. These data demonstrate that punicic acid is inducing apoptosis as part of its cytotoxic effect.
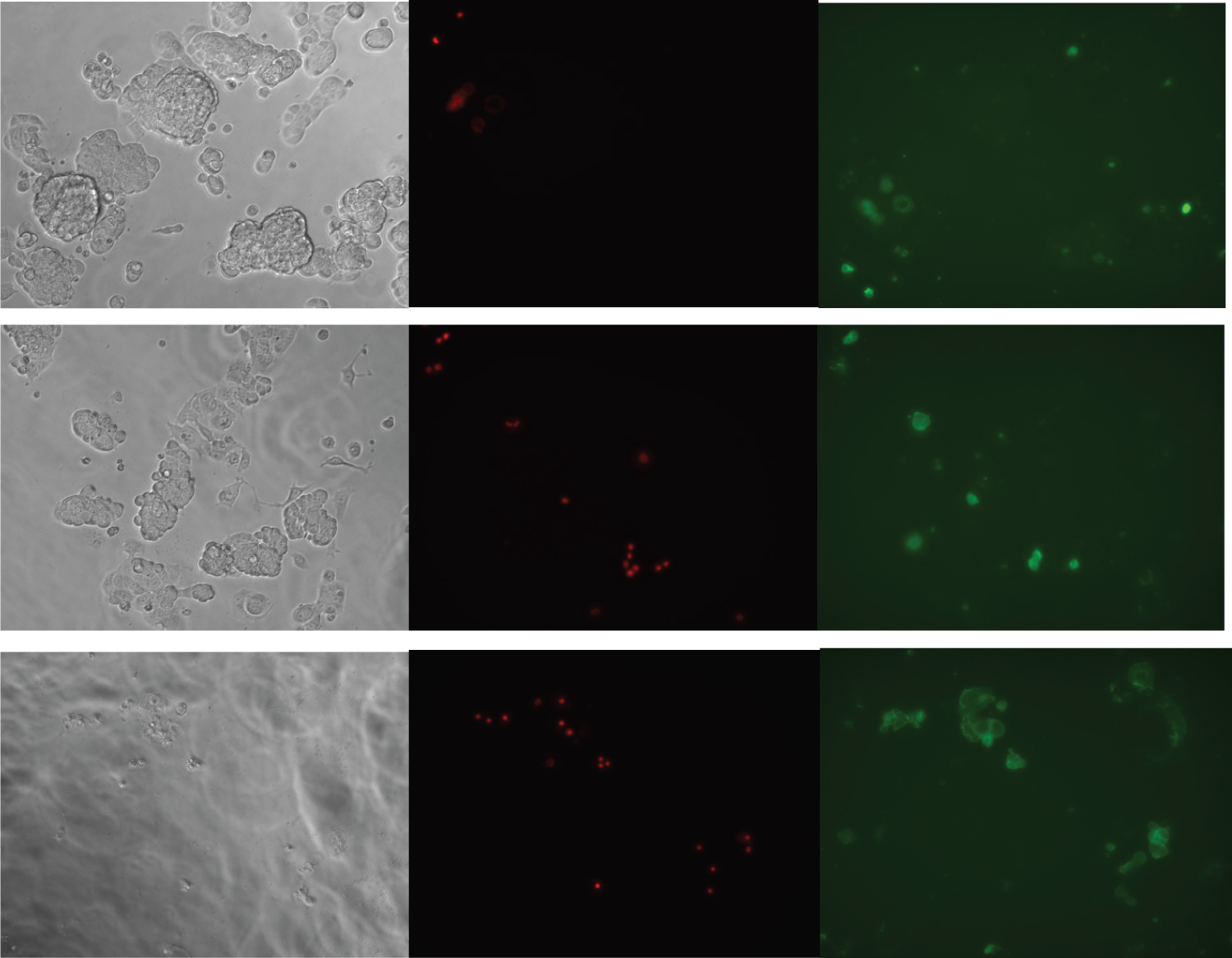 Figure 4: Induction of Apoptosis by Punicic Acid.
Figure 4: Induction of Apoptosis by Punicic Acid.
MCF-7 cells were treated with ethanol control (top), 10 ug/ml punicic acid (middle) or 50 ug/ml punicic acid (bottom) for 48 hours and assayed for apoptosis. Cells were imaged with the Keyence microscope at 200 X using phase contrast (left), red fluorescence to detect Propidium Iodide (center), and green fluorescence to detect Annexin V (right). Cells showing nuclear red staining and membrane-localized green staining are apoptotic.
View Figure 4
While the cytotoxic and apoptotic effect of punicic acid on MCF-7 cells was clear, its effect on gene expression was of interest. Since MCF-7 cells overexpress the peroxiredoxin antioxidant genes, we wanted to know whether these genes are up- or down-regulated in response to punicic acid prior to initiation of cell death. We measured the mRNA expression of Prdx1, Prdx2, Prdx3 and Prdx4 after 24 hours of treatment. As shown in Figure 5, punicic acid led to an increase in Prdx1, Prdx2, Prdx3 and Prdx4, with statistically significant increases found for Prdx1 and Prdx4.
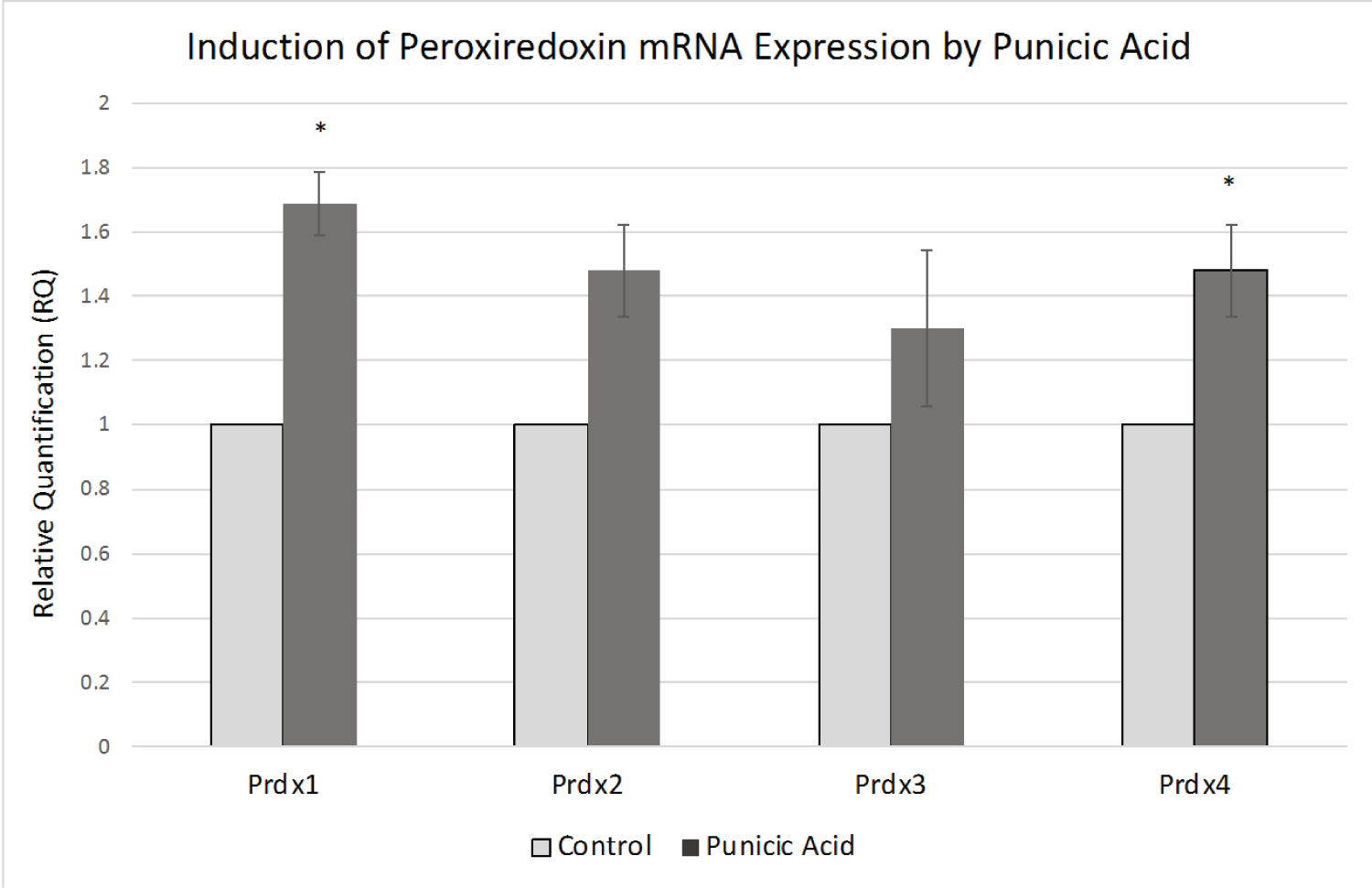 Figure 5: Effect of Punicic Acid on Peroxiredoxin mRNA Expression.
Figure 5: Effect of Punicic Acid on Peroxiredoxin mRNA Expression.
MCF-7 cells were treated with ethanol control or 10 ug/ml punicic acid for 24 hours and Prdx mRNA expression was measured by real time PCR in triplicate reactions, normalized against 18s expression. Control levels for each Prdx were standardized to a value of 1, and average RQ values in control and treated cells are shown (+/- stdev). *p < 0.05.
View Figure 5
The results of our study demonstrate antiproliferative and pro-apoptotic effects of punicic acid on MCF-7 breast cancer cells. We showed a time-dependent and dose-dependent effect on cytotoxicity in this cell line, and also additionally demonstrated induction of Peroxiredoxin gene expression within 24 hours of treatment. These findings are consistent with previous studies on pomegranate seed oil (PSO) and its effect on various cancers including breast cancer [10-15], suggesting a strong role for punicic acid in the PSO-induced effects in these cell lines.
Our data are also consistent with an earlier study showing punicic acid-induced cytotoxicity in two other breast cancer cell lines, MDA-MB-231 cells and the estrogen sensitive MDA-ERalpha7 cell line [17]. While the precise way in which punicic acid elicited its cytotoxic effects in these cells is unknown, Grossman, et al. showed that punicic acid-induced inhibition of proliferation was dependent on lipid peroxidation and activation of the PKC pathway [17]. A very recent study also demonstrated that punicic-acid treated carcinoma cells exhibited ferroptopic cell death, further supporting an important role for lipid peroxides in the mechanism of action for punicic acid-induced cytotoxicity [19]. Understanding the particular signal transduction pathways and cellular processes by which punicic acid mediates its effects will be a critical area of future investigation.
Aberrant signaling from anti-cancer agents often leads to modification of antioxidant gene expression. Peroxiredoxins are thiol-specific antioxidant proteins that reduce a range of cellular peroxides in many cell types [20-22]. This gene family has been demonstrated to play a protective role in several cancers including breast cancer [23,24]. Our current demonstration that Peroxiredoxins are induced in MCF-7 cells by punicic acid treatment is consistent with the role of these cellular antioxidants in cancer cell survival. We previously reported that MCF-7 cells have elevated levels of Peroxiredoxins 1-5 [25,26] and that suppression of several Peroxiredoxins leads to decreased MCF-7 cell survival, and increased resistance to doxorubicin-induced cell death [27]. We also reported upregulation of Peroxiredoxins in tumor tissue of breast cancer patients [28], suggesting that this upregulation is biologically reproducible in-patient tumors.
Over the last few years, the therapeutic potential of antioxidants for cancer treatment has been examined in closer detail, and the role of dietary antioxidants for breast cancer treatment is gaining momentum [29]. Our findings suggest that punicic acid may be considered as a possible new candidate for natural antioxidant treatment of breast cancer. As such, it will be important to further study its mechanism of action in these and other breast cancer cell lines.