Ecological risk assessments (ERAs) for contaminated terrestrial sites regularly include mammals as receptors-of-concern whose home ranges dwarf the sites by tens of times. As a consequence, opportunities to sustain chemical exposures of concern are greatly limited. In having notably miniscule densities as well, wide-ranging mammals are poorly selected species to evaluate because true populations are effectively non-existent at sites. Collectively, the case can be made that wide-ranging mammals are spatially irrelevant at sites to a point where ERAs can be streamlined through their non-consideration. Paralleling work done with the White-tailed deer (Odocoileus virginianus), we used global positioning system (GPS) technology to determine whether or not the only other wide-ranging mammal routinely evaluated in terrestrial ERAs aside from deer, the fox (here, the gray fox, Urocyon cinereoargenteus), is spatially relevant for assessments. Wearing programmable GPS collars, precise fox locations were recorded over periods of either six months or one year as the animals roamed freely. The plotted locations were used in a surrogate manner to establish the percentage of a fox's time spent in land parcels of the size of common hazardous waste sites (i.e., 1, 5, 10 and 20 acres). Fox were observed to be absent from their most utilized parcels more than 80% of the time, and numerous site-size parcels were hardly used. Additionally, the probability of a most utilized 20-acre parcel happening to be a contaminated site was conservatively estimated to be less than 2%. These findings of spatial irrelevance facilitated a systematic review of the worthiness in selecting mammals of any type for evaluation in terrestrial ERAs. For a variety of reasons ranging from the absence of reporting of decimated populations at contaminated sites, to species having extremely low densities, the review found that mammal inclusion in ERAs can be dispensed with, a challenging notion for ERA practitioners to concede. Recognizing the non-need for mammal inclusion in ERAs however, can allow for ERA process to focus on other species and site features more deserving of review.
Ecological risk assessment, Gray fox, Home range, Mammals, Receptor of concern, Spatial relevance, Total area utilized, Urocyon cinereoargenteus
The overwhelming majority of contaminated terrestrial sites that submit to ecological risk assessment (ERA), the focus of this paper, is relatively small. Typical hazardous waste sites extend to only a handful or two of acres or a bit more. This reality holds true even for National Priorities List (NPL) sites of the Environmental Protection Agency's Superfund program, populated with what are reputedly the nation's worst chemical actors. Sixty percent of NPL sites occupy 20 acres or less [1], with this statistic exaggerated in part, because it reflects groundwater sites that have no terrestrial ERA tie-ins, but have plumes that frequently extend for one or more miles [2]. Relatively small chemically contaminated terrestrial sites present a recurring difficulty for ERAs when these evaluations commonly incorporate wide-ranging mammals (e.g., deer, bear). Because these animals traverse terrain that extends far beyond a site's boundaries, their potential for developing toxicological responses of concern from their small-site exposures is drastically reduced, a point that ecological risk assessors may overlook. When ERAs for relatively small sites nevertheless regularly include larger, wide-ranging mammals, the prominent receptor-of-concern selection criterion of having a high degree of site fidelity [3-5] is not being applied. While ERA guidance and other relevant literature do not define a high degree of site fidelity within a bright-line context, it is nevertheless understood that minimal amounts of time spent at a site (e.g., 10%) do not satisfy the intent of receptor-of-concern selection. Thus, the inclusion within a site's developed receptor-of-concern list, of an animal that is absent from a site some 90% of the time, is not easily defended. At a minimum, the inclusion of any receptor (i.e., not only a mammal) with strikingly minimal site exposures, overlooks two critical phenomena. There is first the receptor's potential to clear itself of any site-derived toxins it may have assimilated. Additionally, there is a heightened prospect that a receptor's potentially harmful chemical exposures accrue to some other location. Despite these realities, ERAs are not uncommonly marked by the inclusion of unrealistic mammalian exposure scenarios, such as considering a Louisiana black bear (Ursus americanus luteolus) spending 100% of its time at a one-acre site. Only a tethered or otherwise unnaturally confined bear would actualize such a scenario. Importantly, capitalizing on opportunities to demonstrate the spatial irrelevance of wide-ranging mammals can lead to the streamlining of ERAs, whereby only truly site-representative mammals are the ones retained for evaluation.
To date, demonstrating the spatial irrelevance of routinely assessed wide-ranging mammals at relatively small sites using actual exposure data from the field has been limited to research with the White-tailed deer (Odocoileus virginianus) [2,6,7]. With an average home range of 640 acres [8,9], this species was shown to spend only minimal amounts of time at parcels within its total area utilized (TAU; the territory encompassing all of an animal's recorded locations pursuant to it being allowed to freely roam over many months) that approximate the size of the majority of hazardous waste sites [10]. Conventionally, animal tracking collars employing global positioning system (GPS) technology have weighed too much to be deployed on mammals smaller than deer. The relatively recent design of smaller light-weight collars however, now make spatial movement tracking of some smaller mammals possible [11]. The present study represents one of the first efforts involving the use of GPS collars to evaluate the spatial movements of fox, specifically the gray fox (Urocyon cinereoargenteus). For all intents and purposes, the fox is the only other larger mammal that is commonly selected as a receptor-of-concern in desktop ERAs for terrestrial sites. Its low density aside (an additional factor often contraindicating wide-ranging mammal inclusion in ERAs; discussed further below), with a reported home range in excess of 400 acres [12-14], gray fox, similar to White-tailed deer, would be expected to be spatially and thereby ERA-irrelevant in a typical contaminated site setting. To the extent that gray fox might nevertheless remain spatially relevant for ERA in some way, actual space utilization data for this species (i.e., measures of the degree of contact made with potentially contaminated land parcels) was sought. To the extent that gray fox space utilization data would support larger mammals being inappropriate for ERAs, this study's overarching aim was to assemble a cogent argument for mammals overall, failing to be appropriate for inclusion in ERAs due to varied reasons.
The study area known as Ichauway, in southwestern GA (Baker County; (Figure 1), sits within approximately 12,000 hectares (ha; approximately 30,000 acres) of almost exclusively undisturbed land in the Southeastern Coastal Plain, primarily managed to maintain and restore the longleaf pine (Pinus palustris)-wiregrass (Aristida beyrichiana) ecosystem. The entirety of the study area was conducive to gray fox activity, with land use that included 7,250 ha of longleaf pine forest, and the remaining area primarily consisting of slash (P. elliottii) and loblolly pine (P. taeda) forests, mixed pine-hardwood forests, and lowland hardwood hammocks [15]. Pine forests were characterized by an open canopy, a sparse midstory, and a dense herbaceous understory. Ichauway's surrounding lands were dominated by large center pivot agricultural fields with hardwood forests, pine forests, mixed pine-hardwood forests, pasture/food plots, with residential areas interspersed throughout.
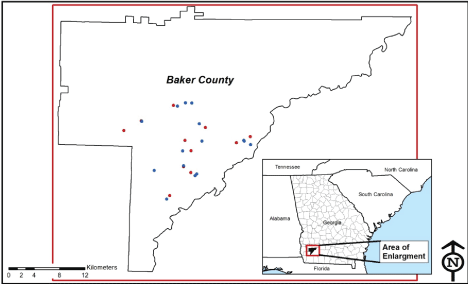 Figure 1: Collaring locations of the study's 26 gray fox in Ichauway, Baker County, GA. Blue dots indicate males; red dots indicate females.
View Figure 1
Figure 1: Collaring locations of the study's 26 gray fox in Ichauway, Baker County, GA. Blue dots indicate males; red dots indicate females.
View Figure 1
Fifteen male and eleven female gray fox (M1-M15 and F1-F11, respectively) were captured with foothold traps in numerous Ichauway locations in 2014 and 2015. After sex, body weight, and general health condition of the fox were recorded, the restrained fox were fitted with GPS collars (Advanced Telemetry Systems, Isanti, Minnesota, USA), each set with a unique VHF frequency. Collars were initially programmed to record precise latitude/longitude (lat/long) locations 7-8 times per day, a rate that would ensure sampling for a full calendar year before battery life would wane. Due to collar malfunctions that occurred in the study's first half year, wherein lat/long locations were found to have not been recorded over periods of months, all collars subsequently deployed were programmed to record locations over periods of six months only. Switching to the shorter tracking period allowed for an augmented sampling rate of approximately 12-16 locations per day. All collared animals were released for their tracking period at their point of capture. Collars were retrieved in one of three ways: utilizing the collar's drop-off function, homing in on the VHF signal in the case of an animal that had died, and via animal recapture. For a limited number of animals, the collar type used allowed for lat/long locations to be remotely downloaded, in which case collars did not have to be retrieved. Table 1 provides animal identification and relevant GPS tracking information (e.g., location sampling rates, total recorded locations) for the study animals. Figure 1 depicts the animal-specific capture/collaring locations. All animal handling and care procedures employed in the study were prior-approved by the U.S. Army's Animal Care and Use Review Office, Office of Research Protections, and by the University of Georgia's Institutional Animal Care and Use Committee.
Table 1: Animal size and tracking statistics summary for adult gray fox (Urocyon cinereoargenteus). View Table 1
Lat/long locations were analyzed using ArcMap, version 10.4 [16]. After initially plotting the constellation of collected locations for each fox, the TAU, as the area encapsulating the totality of space utilized by a fox over the tracking period, was constructed by connecting outermost plotted points. The constellations were then overlain on a series of grids, each made up of cells of a uniform size (e.g., 10 acres), reflecting the sizes of the contaminated sites that most commonly submit to ERAs across numerous regulatory programs. The number of an animal's recorded locations that corresponded to each cell for grids at the 1, 5, 10, and 20-acre scales were tallied, and the percentage of an animal's total number of lat/long locations that coincided with each individual cell of a grid at each scale, was computed. Following Tannenbaum, et al. 2013 [10], the operating assumption was that the percentage of the total number of an animal's recorded locations falling within a given cell would serve as a reasonable approximation of the percentage of that animal's time spent within it.
An additional computation gauged the likelihood of a fox's most utilized cell within its TAU coinciding with a theoretical contaminated site. The analysis recognized that contaminated sites in the U.S., even where those of all regulatory agencies with cleanup programs are combined, do not frequently occur in a spatial framework. For each study animal, the computation took the form of a ratio, with the minimum number of contaminated sites there could reasonably be within a TAU as the numerator, and the number of (20-acre) cells within the TAU that had documented fox utilization, however minimal, as the denominator.
The total number of lat/long locations supporting the spatial relevance/irrelevance analysis approached 2000 in the case of males and exceeded 2,500 for females (Table 1). The greater number of locations for the females primarily reflected their fortuitously having been tracked an average of six weeks more than males, although the higher mean sampling rate for females was also a contributing factor. The greater female sampling rate, although not statistically higher, was also fortuitous; relatively more females than males were collared after it was decided to track animals for a half-year instead of for a full year. Animal F1 was the only fox to have been recaptured and collared for a second period of tracking. As a consequence, it bears the study's overall highest tracking day count, a figure more than double that of the next highest tracked female. Of note, even when F1 is removed from the dataset, the mean tracking day figure for females remains greater than that for males, although not statistically so.
For all but two animals, the pattern of recorded lat/long locations was notably clustered with a minimal number of locations distanced from the cluster by 1-2 miles (Figure 2A). One male and one female however, demonstrated a noticeably different spatial movement pattern, with approximately 14% of locations apart from the main cluster, and with some of those points as much as 14 miles away from a cluster's center (Figure 2B). A review of the chronology of the recorded locations for these animals reveals that even the most outlying points were legitimate, i.e., collar malfunction was not responsible for the atypical spatial movement patterns observed.
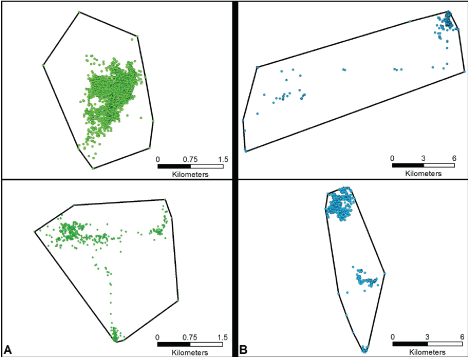 Figure 2: Two types of location (dot) cluster patterns. Panel A demonstrates a gray fox having a dominant area of activity with a lesser degree of points that are relatively short-distanced from that area. Panel B demonstrates a gray fox having a dominant area of activity, and a minimum number of points considerably removed from it. Note that the (kilometer) scalars are different for the two panels.
View Figure 2
Figure 2: Two types of location (dot) cluster patterns. Panel A demonstrates a gray fox having a dominant area of activity with a lesser degree of points that are relatively short-distanced from that area. Panel B demonstrates a gray fox having a dominant area of activity, and a minimum number of points considerably removed from it. Note that the (kilometer) scalars are different for the two panels.
View Figure 2
The four scales of spatial analysis considered are illustrated in Figure 3. The boundary line connecting the outermost recorded lat/long locations, creates the TAU. As the TAU grid size is increased from the one-acre cell size to the 20-acre cell size, the number of cells is, of course, reduced (here, from 1,607 to 100), while the tallies of recorded locations that occur within cells, shift upward accordingly. As is evident, as smaller cells within the TAU coalesce to form larger ones when going from smaller to larger scales, the relative position of the area generally used most by a given fox (i.e., the area where the most utilized cells occur) remains the same.
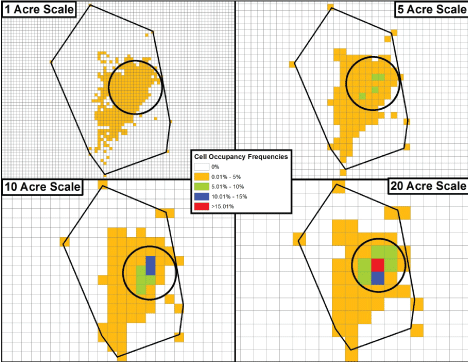 Figure 3: The four scales of spatial analysis for a TAU (the polygon's black outline) are displayed for one of the study animals. Despite the uniform cell size being increased, the relative position of the most highly used portion of the TAU (the circled area) remains the same.
View Figure 3
Figure 3: The four scales of spatial analysis for a TAU (the polygon's black outline) are displayed for one of the study animals. Despite the uniform cell size being increased, the relative position of the most highly used portion of the TAU (the circled area) remains the same.
View Figure 3
The highest cell utilizations for all study animals at each of the four scales are presented in Table 2. As an overall observation for fox at the individual level of analysis, a female nominally (but not statistically) had the highest cell occupancy at each scale. Specifically, where the single most utilized cell overall was considered (the study's '1-cell basis'), F9 had a higher occupancy at the 1, 5, and 10-acre scales over the male with the highest occupancy in each of those cases (M12). At the 20-acre scale, F7 had a higher occupancy than M11, the male with the highest cell occupancy at that scale. At each scale as well, females had higher mean cell utilizations than males with both the '1-cell basis' and the 'top 3-cell basis', the latter an average of an animal's three most utilized cells. While not critical to the overall analysis, there was a singular instance of a statistically higher cell occupancy (p < 0.054) for females, this at the 10-acre scale, where the top 3-cell basis was considered. Importantly, even when using the more conservative 1-cell basis, the highest overall mean cell occupancy figure, for females at the 20-acre scale, was only 24.66% (see Discussion).
Table 2: Highest cell utilization figures for gray fox at different spatial scales (numbers shown are % of total recorded locations). View Table 2
Information to be collectively considered regarding the spatial relevance of fox at typical (i.e., Superfund-type) contaminated sites is presented in Table 3 and Table 4, and in Figure 4, Figure 5 and Figure 6. Table 3 indicates that at any scale, considerable portions of a fox's TAU are often not contacted, and also, greater TAU portions are utilized as scale increases. Figure 4 and Figure 5, for males and females respectively, indicate the frequencies with which the constructed cells at the highest scale considered (20-acres) were utilized. As is evident, the overwhelming majority of cells were utilized no more than 5% of the time in every instance. With regard to the most utilized cells at this scale, there are eight instances for males and three instances for females where only a singular cell had a utilization frequency that exceeded 15%. An additional observation was that only one male (M7) and one female (F8) had a maximum of three cells with greater than 15% utilization.
Table 3: Numbers of uniform-size cells in TAUs and the percentage of cells observed to have been utilized. View Table 3
Table 4: Estimated probabilities (%) of most utilized spaces also being contaminated sites. View Table 4
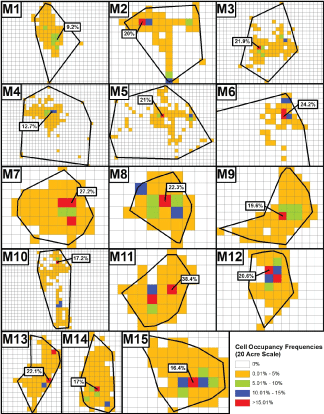 Figure 4: Locations of the single most utilized 20-acre cells within the TAUs of each male gray fox (M1-M15) and the percentage of the animals' time spent within them. Plots are not uniformly sized. Due to M6's considerably large TAU, only a portion of it is shown. Note that none of M6's hidden cells had occupancy frequencies greater than 5%.
View Figure 4
Figure 4: Locations of the single most utilized 20-acre cells within the TAUs of each male gray fox (M1-M15) and the percentage of the animals' time spent within them. Plots are not uniformly sized. Due to M6's considerably large TAU, only a portion of it is shown. Note that none of M6's hidden cells had occupancy frequencies greater than 5%.
View Figure 4
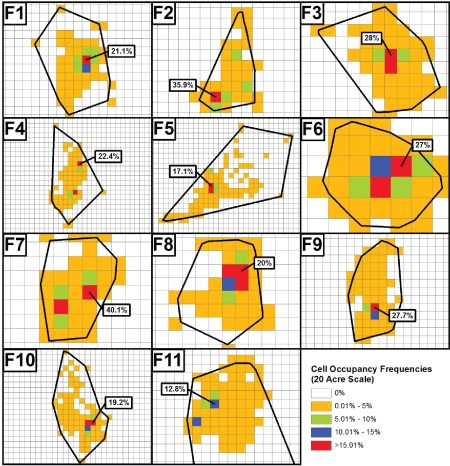 Figure 5: Locations of the single most utilized 20-acre cells within the TAUs of each female gray fox (F1-F11) and the percentage of the animals' time spent within them. Plots are not uniformly sized.
View Figure 5
Figure 5: Locations of the single most utilized 20-acre cells within the TAUs of each female gray fox (F1-F11) and the percentage of the animals' time spent within them. Plots are not uniformly sized.
View Figure 5
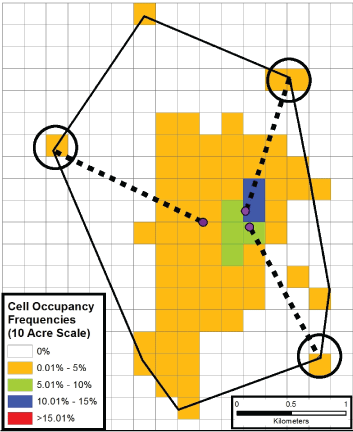 Figure 6: The graphic demonstrates the conservative nature of the estimation of the probability of a most utilized cell also being a contaminated site. En route to occupying the circled outlying cell, a fox had to traverse non-colored cells in order to get there (although non-shaded cells do not reflect lat/long locations having being recorded in them). Purple dots notate a fox's last recorded location before arriving at a circled cell.
View Figure 6
Figure 6: The graphic demonstrates the conservative nature of the estimation of the probability of a most utilized cell also being a contaminated site. En route to occupying the circled outlying cell, a fox had to traverse non-colored cells in order to get there (although non-shaded cells do not reflect lat/long locations having being recorded in them). Purple dots notate a fox's last recorded location before arriving at a circled cell.
View Figure 6
The computed information of Table 4 in conjunction with the highlighted information of Figure 6 broaches the matter of how likely it could be that a fox's most utilized space (in the form of a 20-acre parcel) would also be a contaminated site. Since there could be no more than one contaminated site occurring within a gray fox's TAU (see Discussion), we divided the number 1 by the number of cells that had, even minimally, been contacted (i.e., those shaded). The table indicates that there is no more than a 2% chance of co-occurrence. The information provided by Figure 6, profiling one animal's space utilization at the 10-acre scale of analysis, accentuates this finding. The figure illustrates that certain unshaded cells were undoubtedly contacted (i.e., traversed) in the time delays (of perhaps 3 hours) occurring between consecutive lat/long locations being recorded, for the highlighted locations that were sequentially recorded lie two or more km away from each other. With the legitimized inclusion of at least some additional non-shaded cells in the co-occurrence ratio's denominator, Figure 6 informs that the likelihood of a most utilized cell also being a discrete area of contaminated soil is somewhat less than 2%.
The present study supplies two highly germane pieces of information as corroboration for the assertion that fox are spatially irrelevant at typical sites, and that operationally, selection of the fox as a mammalian receptor-of-concern, is contraindicated. Our data first show that large portions of TAUs are minimally used, and that a fox's most occupied contaminated site-sized parcel, that could in theory be a contaminated one, fails to draw enough of a fox's time to legitimize a chemical exposure health concern. Second, the likelihood of a most occupied parcel also being a contaminated site is miniscule.
Importantly, our data illustrate that fox do not satisfy the prominent receptor-of-concern selection criterion of having a high degree of site fidelity. At the one- and five-acre scales, the mean cell utilizations for the 26 fox we monitored, even where the more conservative '1-cell (utilization) basis' is applied, illustrate that fox do not spend enough time at these areas to matter. Thus, a 15.59% mean cell occupancy for females at the 5-acre scale (Table 2) connotes that some 84% of the time, a gray fox is not utilizing its most preferred location. Cell utilization at the 10 and 20-acre scales communicate the same finding. Further, the computed overall highest mean cell utilization figure of our study (for females at the 20-acre scale, where the '1-cell basis' is applied) informs that some 75% of the time, a fox would not be expected to be within its most favored area. This figure grows to 83% of the time where the more reasonable 'top 3-cell' basis is used. Relevant to this discussion, it is critical to note that a fox is far from being chemically exposed every moment it appears at a contaminated location. Fox do not continuously feed, and they are also not exposed during the hours they sleep (recalling that it is only the dietary ingestion pathway that is evaluated in terrestrial ERA). Although not demonstrated here, had TAUs been divided into uniform cells of 30 and 40 acres, highest cell utilizations would remain essentially the same as those at the 20-acre scale. As explained in part above, as the number of gridded cells of a TAU is decreased (i.e., as cell size is increased), the ratio of the number of cells with relatively higher utilizations to the total number of cells within a TAU, is roughly preserved. Importantly, considering our cell utilization information alone, evaluating fox at typical contaminated sites is contraindicated.
We believe the level of refinement of our spatial movements analysis warrants particular attention. Our data are considerably more robust than what would have been provided with a spatially explicit exposure modeling (SEEM) exercise [17]. Owing to the weaknesses of such modeling, animals are assumed to move across landscapes, guided by variations in habitat quality and foraging areas, and with a new day's foraging activities necessarily linked to the foraging activities of the previous day. Our actual animal movement data indicate land usage patterns that are radically different from such projections. Importantly, a SEEM or other modeling effort would not have been able to depict the actual locations of the parcels contacted, supply the all-critical frequencies with which contact occurred, or establish the sequences with which various locations within a TAU were visited. Thus, a modeling effort could not have approximated the space utilization pattern exhibited in Figure 2B's lower panel.
With regard to the likelihood of a gray fox's most utilized TAU parcel (cell) happening to be contaminated, it is useful to recall that contaminated sites in the U.S., even where those of all regulatory agencies with cleanup programs are combined, do not frequently occur in a spatial framework. This is perhaps best demonstrated by the deployment of Superfund sites in the state of New Jersey, the state with the highest NPL tally, currently at 116. Tens of miles and nearly entire counties separate these contaminated sites. It follows that there would be no more than one contaminated site occurring within a fox's TAU, that for the subject study averaged 3,830 acres in size. More to the point, for the collective area over which our study's 26 fox traveled, estimated to be 429 km2 (165.3 mi2), no more than one contaminated site would be expected.
The conservative nature of the probabilities reported in Table 4 merits review, as follows. Figure 6 illustrates that there are contacted locations within each animal's TAU that are not continuous with the bulk of the TAU portion predominately utilized (i.e., the shaded portions). In the figure's 10-acre scale demonstration however, it is clear that several unshaded cells lying between two consecutively recorded locations must have been contacted, albeit during the 2-3 hours, that on average, transpired between sampling events. For the computation of the probability of a most utilized cell also being a contaminated site then, a universe of cells somewhat greater than just those cells that had some demonstrated utilization should constitute the denominator of the computation. Hence a more accurate probability of a fox's most utilized cell being contaminated, is something less than the maximal 2% figure shown in Table 4. In critical summation, the collective data show that at any scale of analysis, there appears to always be a cell that a given fox uses to some degree more than any other, but with the percentage of the animal's time spent there being rather minimal. With the 20-acre scale of the present study being the most realistic, a legitimate basis for fox inclusion in ERAs is not found; the demonstrated degrees of site exposure are insufficient to generate concerns that fox might succumb from the contaminated soil contact events they might experience. Importantly in this regard, the literature does not describe a phenomenon whereby ecological receptors are lured to areas of soil contamination via olfaction or some other sense, and independent of the contaminant category (i.e., organic or inorganic) present.
We think it noteworthy that the gray fox data highly parallel the White-tail deer data of Tannenbaum, et al. 2013 [10]. The mean percentage of time that deer spent at their most preferred 20-acre parcels was 23.1%, a figure strikingly similar to that computed for the gray fox of this study. Importantly, spatial irrelevance findings for fox are independent of a consideration of a second and undeniable spatial factor at play, namely animal density. Thus, preceding our study, it was generally known that the gray fox has a density that parallels that of its relative, the red fox (Vulpes vulpes) at 0.019 - 0.03 animals/acre [18] or less [8]. These two aspects of the fox's spatial ecology contraindicate its being selected as a mammalian receptor-of-concern for the overwhelming majority of terrestrial sites that submit to ERA, this paper's consideration. In the common case, sites are vastly smaller than the fox's home range, that for our study was estimated to span 368 to 543 acres (data not presented), and sites at best can be expected to house no more than one monogamous pair. Arguably for ERAs, the inclusion of mammal species with animal counts so very low is a wasteful and misleading practice. Regardless of how much time an animal might spend at a site, remediation work to afford protection to just one or two species representatives that contact it, and that do not effectively constitute a population, does not proceed. This is certainly so where special status species are involved [4].
Fox and White-tailed deer are very likely the two most commonly evaluated larger mammals of terrestrial ERAs conducted in the U.S., if not the only two. With information now available to demonstrate that neither of these species is spatially relevant for land parcels that are the size of most contaminated sites, ERA consequences should be considered. A first observation is that the stated intent of mammalian receptor-of-concern selection appears to be commonly corrupted. Receptors-of-concern are selected for the purposes of streamlining ERAs. Guidance and standard practice recognize that numerous species populating a given site need not be independently assessed where one particular species can represent others, provided the biologies for all are similar. Here we are prompted to ask for what similar mammals are gray fox and White-tailed deer ever selected to serve as surrogates in ERAs? If the answer should be those mammals for which there would only be one or two species representatives anticipated present, a basis for assessment is lacking altogether. Further, it is doubtful that there are large omnivorous and herbivorous mammals other than fox and deer, respectively, with notably greater densities than these two species. In support of this, reported densities for mink (Mustela vison) and kit fox (Vulpes macrotis) are 0.04 and 0.001 animals/acre, respectively [8,18-21]. If there should be larger mammals with notably higher densities, we question why fox and deer, with sparse distributions that suggest only singular animal representation or perhaps the presence of no more than one monogamous pair at typical sites, are the ones routinely selected to be surrogates.
With regard to this density discussion, it is particularly relevant to recall that with the fox figures available, it would be quite unlikely that a given fox's most utilized cell would be regularly occupied by a second fox. Additionally, it would be a forced assumption that wherever gray fox occur, each animal's most utilized location happens to be a den situated within a contaminated site, and particularly when a den typically approximates the size of an office cubicle. Our lat/long data that include the corresponding times for all recorded locations, did not illustrate any of the foxes commonly occupying a fixed den-sized location over many days. To the extent that the presence of a stream or other water source might account for greater time spent at the most utilized cells, we used an imagery layer in ArcMap to review these locations. For the most utilized cells of several randomly selected gray fox, we found none that had a stream or other waterbody present.
The notion that no mammals at all might warrant a place in terrestrial ERAs for conventional sites is disconcerting to many ERA practitioners [2]. Finding the notion untenable might first be traceable to miseducation. Ecological risk assessors may believe that regardless of circumstance (e.g., it being known that a species has a minimal site presence), there is a requirement that terrestrial ERAs always include at least one wide-ranging mammal. No such requirement exists, however. Further, independent of the numerous shortcomings of the routinely applied hazard quotient computation and the overall ERA process [22,23], assessors might also find the notion untenable because it suggests that efforts invested to characterize the chemical exposures of mammals are wasted ones [23]. Reasonably though, the data we furnish assert that there is little point to including species that are frequently absent from a site.
With defensible arguments now in place for ERAs excluding wide-ranging/sparsely distributed mammals from consideration, the remaining discussion asks if there might be any mammals at all that are appropriate for inclusion in terrestrial ERAs. A consideration of small rodents (e.g., field mice) and related insectivores (e.g., shrews), receptor-of-concern list fixtures in terrestrial ERAs, is a good starting point. Small rodents will always be spatially relevant because of their home ranges that rarely exceed one acre, and because of their relatively high densities. There are other reasons though, that support their non-selection. First, accounts of depauperate small rodent populations at contaminated sites are lacking [4], and sites with exceedingly high soil contamination have illustrated that reproduction in rodents is unimpaired [24]. This observation continues to fuel a primary tenet of ERA's only direct health status assessment method, Rodent Sperm Analysis [25]. The method understands how the small rodent, in a spatial sense, can serve as a screen for the potential development of contaminant-mediated health effects in larger mammals. The method posits that if maximally exposed rodents (i.e., with an inability to move beyond contaminated surroundings) are not found to be harmed by their chemical-in-soil exposures, wider-ranging mammals with their vastly reduced degrees of site contact, should certainly not be health-challenged by the contaminant exposures they may incur [2,25,26].
An additional reason to dispense with small rodent assessments, although not readily illustrated [4], notes that historically, terrestrial sites have not been remediated for the sole purpose of affording greater protection to this animal group. Further, while some ecological risk assessors may argue that high site fidelities allow small rodents to body-store contaminants that are subsequently transferred to higher carnivorous species to pose potential risks, such reasoning is easily challenged. Because the higher trophic level receptors (e.g., hawk, fox) have relatively huge home and foraging ranges, they would not be regularly (or only) consuming contaminant-bearing rodents of five- or ten-acre parcels and the like to satisfy their dietary needs [2]. It is important to note that other relatively small mammals, aside from those of the Order Rodentia, are also sparsely distributed, and to the point that not enough of them would occur within a 20-acre parcel to pose a concern. As an example, the striped skunk (Mephitis mephitis), with its highest density considered, might have two individuals present within such a space [19].
With wide-ranging mammals seemingly inappropriate for ERA inclusion due to spatial irrelevance, and small rodents and other relatively small mammals inappropriate because of their demonstrated contaminant resiliency, only a handful of mammals remain to consider in an ERA context. It would seem that these too would not be deemed appropriate for ERA inclusion where their distinct biologies are reviewed. The long-tailed weasel (Mustela frenata), a decidedly larger form than most small rodents (generally measuring from 37 to 50 cms in length, not including the tail), is one case in point. Because a weasel weighs as much as a GPS collar, it cannot submit to the spatial movement tracking described in this paper. The weasel though, has a home range of 300 acres, and a density range of 1 animal per 7-40 acres. At its maximum density, a 20-acre parcel (describing again, a very common size of contaminated site) could be expected to support no more than three of these animals, an insufficient number to generate an ecological risk concern. Additionally, it would be unrealistic to suggest that a weasel (and only one weasel) spends perhaps 85% of its time within a 20-acre area of its home range (an area that could be a contaminated site) and spends the remaining 15% of its time occupying the other 280 acres of its home range. Instances of such highly skewed spatio-temporal partitioning are unknown for the mammals conventionally reviewed in ERAs. The commonly ERA-evaluated raccoon (Procyon lotor) bears brief mention here. While this species' density may be several orders of magnitude greater than that of the earlier mentioned kit fox and mink, its home range can often extend to the hundreds of acres [8], thereby aligning more with fox and deer in terms of being highly unlikely to be substantially contacting typical contaminated sites. A final consideration is the badger (Taxidea taxus), a species not often evaluated because it does not occur in all states. A solitary species, it has a reported average density of five animals in 247 acres [27]. For sites that measure less than 50 acres in size, as most do, there would an expectation of no more than one individual present. Here, an additional reason to forego certain mammal assessments at contaminated sites arises; ERAs are supposed to assess populations and not individuals [28], and realistically there would not be a badger population to speak of at a commonplace 1, 5, 10, or 20-acre contaminated site.
With our gray fox spatial movement tracking data being highly corroborative of similar data for the White-tailed deer, including wide-ranging mammals in terrestrial ERAs for typical contaminated sites appears to be effectively without purpose. We believe this conclusion provides a proper springboard for a cautious review of the worthiness of including other mammals as receptors-of-concern in ERAs. For multiple reasons, ranging from species being sparsely distributed to their displaying noteworthy resiliency despite unavoidable chemical exposures incurred, the case for dispensing entirely with mammal assessments is supported. Such streamlining of future ERAs, if implemented, would facilitate placing a heightened focus, where needed, on other species and site features more deserving of review.
We thank the U.S. Army Environmental Command for funding this study. We appreciate the skillful manuscript reviews of Jeffrey Leach and Sherri Hutchens. The authors do not have any conflict of interests to declare.