Vitamin A (retinol) is an essential lipid-soluble vitamin important for a number of biological functions. In mice, reduction of its dietary content by substituting standard rodent chow with a chow containing 5 times less vitamin A and feeding mice for three generations with this modified diet, results in the depletion of all-trans- retinoic acid. In the present study we investigated if the low vitamin A diet affects skeletal muscle function. Mice fed the low vitamin A diet ran significantly less and at a slower speed compared to age matched chow fed mice. The specific force and Ca2+ transients developed upon delivery of a train of pulses, as well as glycogen stores were reduced in muscles from low vitamin A diet fed mice. Furthermore, slow twitch muscles from low vitamin A fed mice fatigued more rapidly and exhibited a higher degree of protein oxidation after electrical stimulation. In summary, these results demonstrate that a chronically low vitamin A diet negatively impacts skeletal muscle function.
Low vitamin A diet, Muscle force, Excitation contraction coupling, Oxidation, Glycogen
CSQ: Calsequestrin; Cav1.1: Alpha 1 Subunit of the Dihydropyridine Receptor; ECC: Excitation Contraction Coupling; EDL: Extensor Digitorum Longus; LVAD: Low Vitamin A Diet; MyHC: Myosin Heavy Chain; RyR: Ryanodine Receptor
Vitamin A is a lipid-soluble anti-oxidant essential vitamin important for a variety of biological functions including eyesight, modulation of energy metabolism, maintenance of soft tissues, mucous membranes and skin as well as in sustaining the immune system [1,2]. Its absence is linked to abnormal fetal development and in rodents its dietary lack causes developmental delays, low body weight and breakdown of skeletal muscle [3]. Although the ergogenic effect of dietary vitamin A supplements and of other anti-oxidants is still a matter of debate [4], a recent study from our laboratory demonstrated that mice over-expressing the retinol dehydrogenase SRP-35 in their skeletal muscles showed enhanced muscle performance [5]. SRP-35 is a 35 kDa membrane bound protein of the sarcoplasmic reticulum (SR) belonging to the DHRS7C [dehydrogenase/reductase (short-chain dehydrogenase/reductase family) member 7C] subfamily [6-8]; metabolizing retinol (Vitamin A) to all-trans-retinaldehyde [8], the substrate for the irreversible oxidative reaction generating all-trans retinoic acid (ATRA). SRP-35 transgenic mice exhibited a number of metabolic changes leading to enhanced running capacity, enhanced glucose uptake, and a stronger activation of PI3K and of mammalian target of rapamycin complex 2 (mTORC2) signaling pathways [5]. Since SRP-35 is located in the SR, these results imply that vitamin A and/or its metabolites may be important locally, generating retinol in the organelle primarily involved in calcium regulation thereby playing an important role in skeletal muscle function.
In the present study we directly investigated the role of vitamin A in skeletal muscle function and excitation contraction coupling (ECC). To this aim, we kept wild type C57/Bl6 mice on a Low Vitamin A Diet (LVAD) for three generations. Rodents vary in their sensitivity to low dietary vitamin A, with different strains of mice being more resistant to low dietary levels than others [9]. Therefore, in order to study the impact of low vitamin A mice need to be fed for at least three generations with a special diet containing 4 IU/g instead of 20 IU vitamin A/g as in the standard rodent chow. This protocol results in the depletion of all-trans-retinoic acid from various tissues including liver, kidney, white adipose tissue and testis [9]. The results of the present investigation demonstrate that a chronically low vitamin A diet negatively impacts skeletal muscle function in vivo and ex vivo; this effect is likely to be the consequence of multiple factors including smaller skeletal muscle glycogen stores and enhanced protein oxidation.
All experiments were conducted according to the Swiss Veterinary Law and institutional guidelines and were approved by the Kantonales Veterinäramt Basel Stadt (Kantonal permits 1728 and 2115). All animals were housed in a temperature-controlled room with a 12 h light-12 h dark cycle and had free access to food and water.
C57Bl/6 male mice were fed a Low Vitamin A Diet (LVAD; 4 UI/g; Scientific Animal food and Engineering, SAFE Labortories, 89290, Augy, France) for 3 generations. Male mice of the third generation were sacrificed at 12 weeks of age [9] and their muscles removed for biochemical and physiological experiment. Age and sex matched C57Bl/6 control mice were fed SAFE105 diet containing 20 UI/g Vitamin A. This diet contains the same components as the LVAD except for the vitamin A content, and will be referred to as �Standard chow diet� throughout the manuscript.
In vivo muscle function was assessed using the voluntary running wheel on 12-weeks-old male mice as previously described [5,10]. Ex vivo muscle function was assessed by subjecting isolated muscles to field stimulation. Briefly, extensor digitorum longus (EDL) and soleus muscles were dissected from 12 weeks-old mice and mounted onto a muscle force transducing setup (Heidelberg Scientific Instruments) as previously described [5,11]. Muscle force was digitized at 4 kHz by using an AD Instrument converter and stimulated with 15 V pulses for 1.0 ms. Tetanus was recorded in response to a train of pulses of 1.0 ms duration delivered at 50/100/150 Hz, and 50/100/120 Hz, for EDL and soleus, respectively for 400 ms and 1100 ms. Specific force was normalized to the muscle cross-sectional area [CSA = wet weight (mg)/length (mm) × 1.06 (density mg/mm3)].
Resistance to fatigue was measured in EDL and soleus muscles stimulated with a train of tetanic stimulation (EDL: 150 Hz, 400 ms duration; Soleus 120 Hz, 400 ms duration) delivered at 0.27 Hz. Maximal specific force (mN/mm2) of the first tetanic contracture is expressed as 100%.
Single fibers were incubated for 10 min at 19 ℃ in Ringer's solution containing 10 µM low affinity calcium indicator Mag-Fluo-4 AM (Thermo Fischer), 50 µM N-benzyl-p-toluene sulfonamide (BTS, Tocris). Fibers were rinsed twice with fresh Tyrode's solution, and measurements were carried out in Tyrode's solution containing 50 µM BTS as described [11]. Measurements were carried out with a Nikon Eclipse inverted fluorescent microscope equipped with a 20x PH1 DL magnification objective. The light signals from a spot of 1 mm diameter of the magnified image of single fibers were converted into electrical signals by a photomultiplier from Myotronic (Heidelberg, Germany) connected to PowerLab Chart 5 analog digital converted. Fluorescent signals were analyzed using PowerLab Chart 5 and OriginPro 8.6.0 softwares. Changes in fluorescence were calculated as ΔF/F0 = (Fmax - Frest)/(Frest). Kinetic parameters were analyzed using Chart 5 software. Fibers were excited at 480 nm and then stimulated either with a single pulse of 50 V of 1 ms duration, or with a train of pulses of 50 V of 1 ms duration delivered at 100 Hz for 300 msec. Fibers were isolated from 3-5 mice and the results were pooled.
Total homogenates from EDL and soleus muscles were prepared as previously described [5,11]. Protein concentration was determined using a Protein Assay Kit II (Bio-Rad Laboratories, 500-0002) and BSA was used as a standard. SDS-PAGE, protein transfer on to nitrocellulose membranes and immunostaining were performed as previously described [5,11], except for parvalbumin and MyHC. For parvalbumin, proteins were separated on a 10% Tris-Tricine gel [12] and subsequently transferred onto nitrocellulose. For separation of MyHC isoforms, high resolution gel electrophoresis was performed [13]. The specifications and dilutions of the primary antibodies used for Western Blotting are listed in Supplementary Table S1: Anti-RyR1 (Cell Signaling, 8153S), anti-Cav1.1 (Santa Cruz sc-8160), anti-Calsequestrin-1 (Sigma, C-0743), anti-SERCA1 (Santa Cruz, sc-8093), anti-SERCA2 (Santa Cruz, sc-8095), anti-JP-45 [14], anti-parvalbumin (Swant, PV25) and anti-MyHC (Millipore, 05-716). Secondary peroxidase conjugates were Protein G-peroxidase (Life Technologies, P21041) and peroxidase-conjugated goat anti-mouse IgG (Sigma, A2304). The immuno-positive bands were visualized by chemiluminescence using the WesternBright ECL-HRP Substrate (Witec AG). Supplementary Figure S1 shows representative immunoblots. Densitometry of the immune-positive bands was carried out using Fusion Solo S (Witec AG).
Supplementary Table 1: Twitch and tetani force developed by EDL and soleus muscles from chow and LVAD fed C57/Bl6 mice. View Supplementary Table 1
 Supplementary Figure 1: Immunoreactivity of antibodies used on western blots. 30 μg proteins in total homogenates of EDL and soleus from chow and LVAD fed mice were separated on 6% (RyR1), 7.5%, 10% or Tris tricine SDS-PAG, blotted onto nitrocellulose and probed with the indicated primary antibodies. Blots were then incubated with peroxidase conjugated anti mouse IgG or peroxidase conjugated protein-G and the bands were visualized by chemiluminescence.
View Supplementary Figure 1
Supplementary Figure 1: Immunoreactivity of antibodies used on western blots. 30 μg proteins in total homogenates of EDL and soleus from chow and LVAD fed mice were separated on 6% (RyR1), 7.5%, 10% or Tris tricine SDS-PAG, blotted onto nitrocellulose and probed with the indicated primary antibodies. Blots were then incubated with peroxidase conjugated anti mouse IgG or peroxidase conjugated protein-G and the bands were visualized by chemiluminescence.
View Supplementary Figure 1
Oxidized proteins present in total muscle homogenates were identified using the Oxyblot protein oxidation detection kit (Millipore S7150) following the manufacturer's recommendations. Briefly, total homogenates of freshly isolated EDLs and soleus muscles were prepared, the carbonyl groups were derivatized and the samples were then snap frozen and stored in liquid nitrogen until used. For experiments on electrically stimulated muscles, isolated EDL and soleus muscles were incubated at 30 ℃ in Krebs-Ringer buffer, continuously gassed with 95% O2 and 5% CO2. Muscles were stimulated with a 300 ms train of action potentials of 1.0 ms duration at a frequency of 150 Hz for EDL muscles and 120 Hz for soleus muscles, by using a stimulator (Myotronic, Heidelberg Germany); the trains of action potentials were delivered for 10 min at a frequency of 0.27 Hz. At the end of the stimulation protocol muscles were processed as described above, snap frozen and stored in liquid nitrogen until used. Proteins were solubilized in cracking buffer (62.5 mM Tris-HCl pH 6.8, 6 M Urea, 2.3% SDS, 5% β-mercaptoethanol, 10% glycerol), 30 µg were separated on a 10% SDS PAGE or 4 µg in a MyHC high resolution acrylamide gel [13], blotted onto nitrocellulose and processed as described in the Oxyblot kit. Immunopositive bands were visualized by chemiluminescence using the WesternBright ECL-HRP Substrate (Witec AG). Densitometry of the immuno-positive bands was carried out using the Fusion Solo S (Witec AG) and the intensity of the DNPH immune-positive bands was normalized for the intensity of MyHC immune-positive band.
Glycogen content was assessed as previously described [5] on snap frozen EDL and soleus muscles using a hexokinase-dependent kit following the manufacturer's instructions (GAHK-20, Sigma-Aldrich; USA).
Statistical analysis was performed using the software OriginPro 8.6.0 (OriginLab Corporation). Comparisons of two groups were performed using the Student's t-test, for groups of three or more comparisons were made using the ANOVA test followed by Tukey's post-hoc test unless otherwise stated. Means were considered statistically significant when p values were < 0.05. All figures were created using Adobe Photoshop CS6 or R Studio (version 0.99.891 or newer). For non-parametric results, the Mann-Whitney test was used.
In a previous study we showed that mice over-expressing SRP-35 an enzyme involved in the conversion of retinol (vitamin A) to all-trans-retinaldehyde, exhibited enhanced skeletal muscle performance in vivo [5]. In order to demonstrate that this effect is causally linked to vitamin A we placed wild type C57/Bl6 mice on a low vitamin A diet for 3 generations and compared the skeletal muscle function of the LVAD fed mice to that of chow fed mice. In vivo muscle function was investigated using the voluntary running wheel and ex vivo function was investigated measuring electrically evoked force generation. Figure 1a shows that LVAD fed mice (open circles) ran significantly less (27 km less after 21 days; p < 0.05, ANOVA. Left panel) and at a slower speed (right panel) compared to age matched chow fed mice (closed circles). The average speed of chow fed mice was 0.89 ± 0.07 km/h (n = 16) and that of LVAD fed mice was 0.76 ± 0.08 km/h (n = 16) (p < 0.05 ANOVA). The shorter running distance was not due to the LVAD fed mice spending less time on the running wheel as the mean time spent by the LVAD fed mice on the running wheel was 3.9 ± 1.5 hours/day, a value not different from that of chow fed mice (4.1 ± 1.4 hours/day). The reduced running capacity is not linked to changes in fiber type composition since the myosin heavy chain (MyHC) isoform content of extensor digitorum longus (EDL) and soleus muscles from chow and LVAD fed mice is similar (Figure 1b). Additionally, the reduced running capacity was not caused by an overall decrease in muscle mass as the weight of chow and LVAD fed mice was similar (Supplementary Figure S2), nor to changes in the weight of fast and slow twitch muscles (Supplementary Table S2), nor in the content of the main proteins involved in ECC. In fact, the content of ryanodine receptor 1 (RyR1), the alpha 1 subunit of the dihydropyridine receptor (Cav1.1), calsequestrin 1 (CSQ1), JP-45, and SERCA2 was similar in EDL and soleus muscles from chow and LVAD fed mice (Figure 1c). Interestingly, the parvalbumin content in total homogenates of EDLs from LVAD was 73.4% ± 10.3 (p < 0.05, Student t test) of that in EDLs from chow fed mice. In addition, SERCA1 content in total homogenates of soleus muscles from LVAD the was significantly higher (193.9% ± 55.4 p < 0.05, Student t test), than that of chow fed mice (for whole uncropped representative blots, see Supplementary Figure S1).
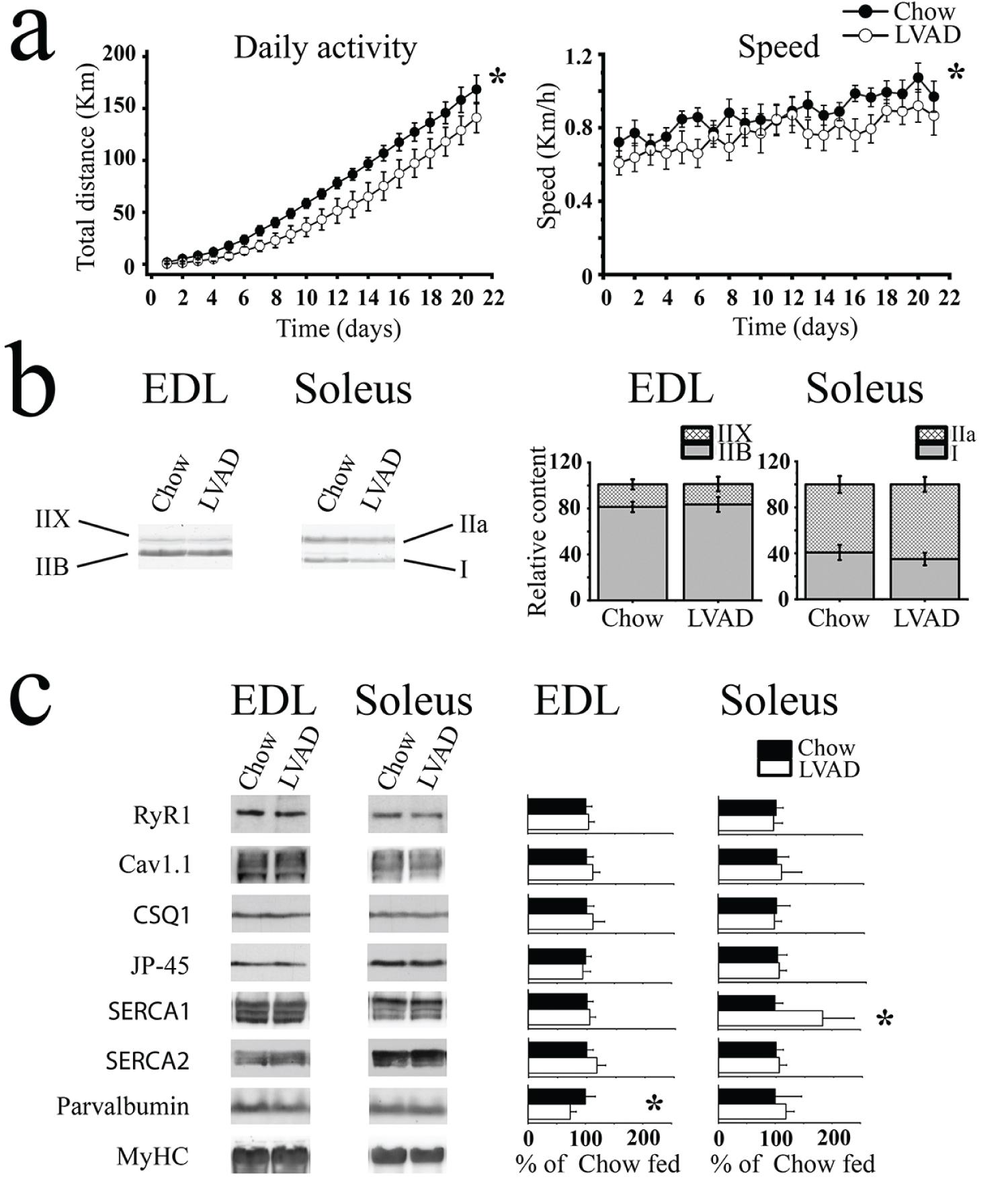 Figure 1: Spontaneous activity is decreased in LVAD fed mice but this is not caused by altered content of RyR1 and Cav1.1. (a) Spontaneous daily activity (left) and mean running speed (right) of chow few (black symbols; n = 16) LVAD fed mice (empty symbols; n = 16) mice. Each point represents the mean (± S.D.) distance run over a period of three weeks *p < 0.05 ANOVA; (b) Representative Coomassie blue stained high-resolution gel showing that there are no differences in MyHC isoform content between chow and LVAD fed mice. The histograms on the right show the mean (± S.D., n = 5) relative expression of each MyHC isoform relative to the MyHC total content; (c) Western blot analysis of total muscle homogenates. Left panels, representative immunopositive bands. Right panel, bar graphs showing mean (± S.D., n = 4-6) relative intensity of the immunoreactive band in muscle homogenates from LVAD fed mice. Homogenates were prepared from 4-6 chow fed and 4-6 LVAD fed mice. The mean intensity of the immunopositive band in chow fed mice was set to 100%. Proteins were separated on 6% (RyR1, Cav1.1), 10% (SERCA1, SERCA2, JP-45, CSQ1) or Tris-Tricine gels (parvalbumin) SDS-PAGE.*p < 0.05 Student's t test.
View Figure 1
Figure 1: Spontaneous activity is decreased in LVAD fed mice but this is not caused by altered content of RyR1 and Cav1.1. (a) Spontaneous daily activity (left) and mean running speed (right) of chow few (black symbols; n = 16) LVAD fed mice (empty symbols; n = 16) mice. Each point represents the mean (± S.D.) distance run over a period of three weeks *p < 0.05 ANOVA; (b) Representative Coomassie blue stained high-resolution gel showing that there are no differences in MyHC isoform content between chow and LVAD fed mice. The histograms on the right show the mean (± S.D., n = 5) relative expression of each MyHC isoform relative to the MyHC total content; (c) Western blot analysis of total muscle homogenates. Left panels, representative immunopositive bands. Right panel, bar graphs showing mean (± S.D., n = 4-6) relative intensity of the immunoreactive band in muscle homogenates from LVAD fed mice. Homogenates were prepared from 4-6 chow fed and 4-6 LVAD fed mice. The mean intensity of the immunopositive band in chow fed mice was set to 100%. Proteins were separated on 6% (RyR1, Cav1.1), 10% (SERCA1, SERCA2, JP-45, CSQ1) or Tris-Tricine gels (parvalbumin) SDS-PAGE.*p < 0.05 Student's t test.
View Figure 1
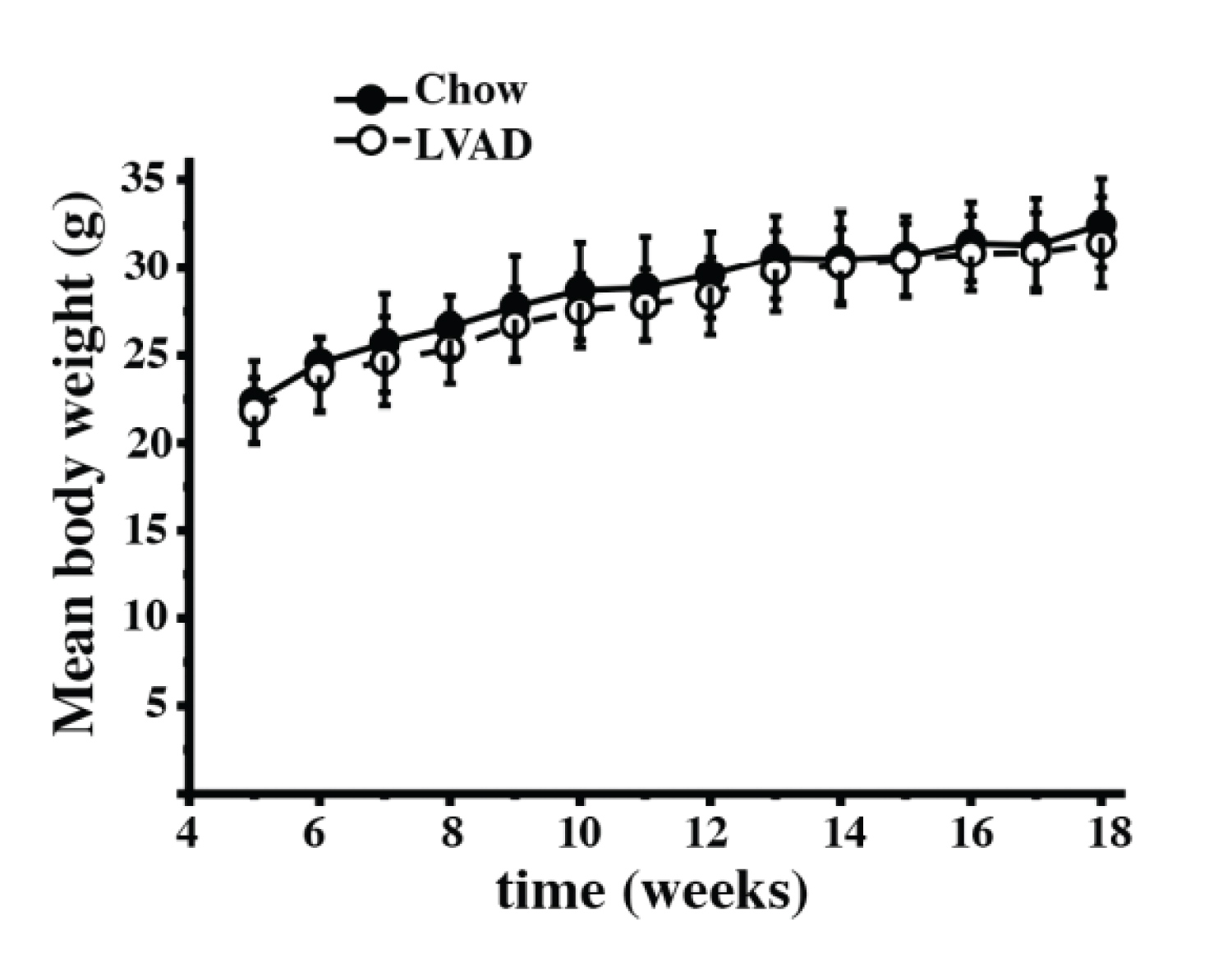 Supplementary Figure 2: Growth curves of Chow and LVAD fed mice. Male mice were weighed once a week starting from week 5 of age. No difference in weight between Chow fed (closed circles) and LVAD fed (empty circles) was observed. Each symbol represents the mean (± S.D.) weight in grams of n = 8 (Chow fed) and n = 9 (LVAD) fed mice.
View Supplementary Figure 2
Supplementary Figure 2: Growth curves of Chow and LVAD fed mice. Male mice were weighed once a week starting from week 5 of age. No difference in weight between Chow fed (closed circles) and LVAD fed (empty circles) was observed. Each symbol represents the mean (± S.D.) weight in grams of n = 8 (Chow fed) and n = 9 (LVAD) fed mice.
View Supplementary Figure 2
Supplementary Table 2: Analysis of electrically evoked calcium transients in EDL and soleus muscles from chow and LVAD fed C57/Bl6 mice. View Supplementary Table 2
We also performed ex vivo force measurements on isolated fast and slow twitch muscles. EDL muscles were apparently more sensitive to the effects of low vitamin A (Figure 2). Indeed, the force developed upon delivery of a single action potential (twitch force, Figure 2a and Figure 2c) or by a train of pulses of 1.0 ms duration delivered at frequencies of 50, 100 and 150 Hz (tetani, Figure 2b and Figure 2c) was significantly reduced in EDLs from the LVAD mice. The average (mean ± S.D.) specific twitch peak force induced by a single action potential in EDL from chow fed and LVAD mice was 99.6 ± 15.3 mN/mm2 (n = 7), vs. 77.6 ± 15.1 mN/mm2 (n = 10), respectively (p < 0.05 Mann-Whitney test. Figure 2c and Supplementary Table S2). Similarly, the specific tetanic force at all tested frequencies was decreased in EDLs (Figure 2c, left panel and Supplementary Table S2) from LVAD mice compared to that of chow fed mice. The twitch force developed by soleus muscles was not affected by the LVAD (twitch force was 48.71 mN/mm2, n = 7 vs. 42.2 ± 12.4, mN/mm2, n = 9, for chow and LVAD fed mice, respectively. Figure 2a), but at high frequency tetanic stimulation (that is at 100 and 120 Hz. Figure 2b and Figure 2c) the specific force was also decreased in soleus muscles from the LVAD fed mice (Figure 2c right panel and Supplementary Table S2).
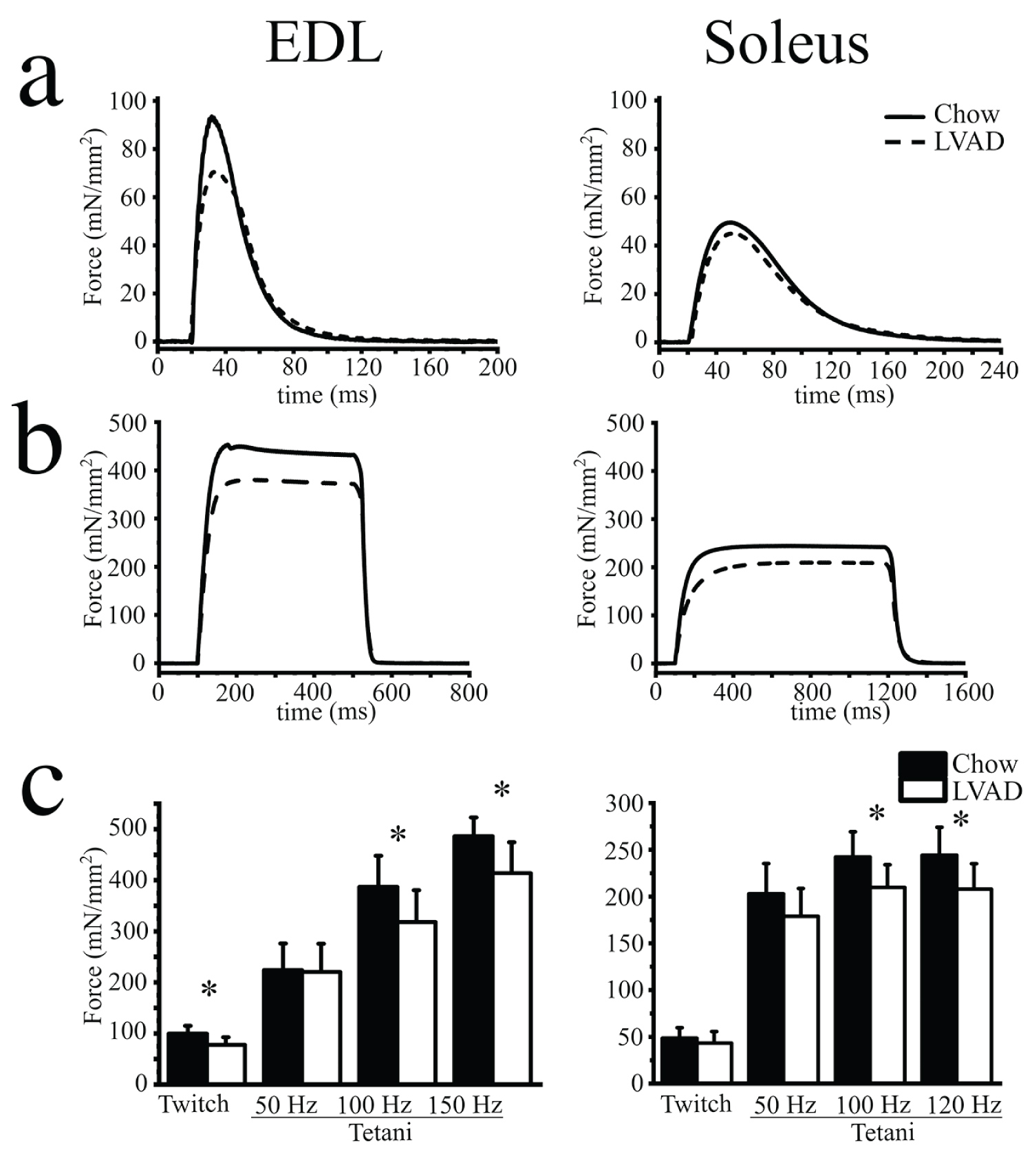 Figure 2: Mechanical properties of isolated EDL and soleus muscles from chow and LVAD fed mice. Representative traces showing force developed by EDL (left) and soleus (right) muscles isolated from chow fed (continuous trace) and LVAD fed (dashed trace) mice. (a) Twitch force was stimulated by a 15 V pulse of 1.0 ms duration; (b) Maximal tetanic force was induced by a train of pulses delivered at 150 and 120 Hz for EDL and soleus, respectively; (c) Mean (± S.D.) specific force (mN/mm2) developed after twitch and tetanic stimulation at the indicated frequencies. Experiments were performed on muscles isolated from n = 7 chow fed and n = 9 LVAD fed mice. * p < 0.05 Mann-Whitney test.
View Figure 2
Figure 2: Mechanical properties of isolated EDL and soleus muscles from chow and LVAD fed mice. Representative traces showing force developed by EDL (left) and soleus (right) muscles isolated from chow fed (continuous trace) and LVAD fed (dashed trace) mice. (a) Twitch force was stimulated by a 15 V pulse of 1.0 ms duration; (b) Maximal tetanic force was induced by a train of pulses delivered at 150 and 120 Hz for EDL and soleus, respectively; (c) Mean (± S.D.) specific force (mN/mm2) developed after twitch and tetanic stimulation at the indicated frequencies. Experiments were performed on muscles isolated from n = 7 chow fed and n = 9 LVAD fed mice. * p < 0.05 Mann-Whitney test.
View Figure 2
In order to determine if the changes in the mechanical properties are caused by alterations of ECC we analyzed electrically evoked Ca2+ transients in isolated muscle fibers (Figure 3). In the presence of 1.8 mM Ca2+ in the extracellular solution, the averaged peak Ca2+ transient induced by a single action potential in EDL fibers from LVAD fed mice was decreased by 16% from a mean (± S.D.) ∆F/F0 of 1.48 ± 0.21 in chow fed mice to 1.24 ± 0.19 in LVAD fed mice (p < 0.05 Mann-Whitney test) (Figure 3a and Supplementary Table S3). The kinetics of the Ca2+ transients were decreased and the Time to peak (TTP) was reduced from 1.46 ± 0.25 ms in chow fed to 1.34 ± 0.20 ms in LVAD fed mice (p < 0.05 Mann-Whitney test) (Supplementary Table S3). In contrast, the peak Ca2+ transients elicited by twitch stimulation of isolated soleus fibers were similar in chow and LVAD fed mice. The summation of Ca2+ transient peaks evoked by a train of pulses delivered at 100 Hz in the presence of 1.8 mM Ca2+ in the extracellular solution in EDL fibers from LVAD mice (Figure 3a) was decreased from a mean (± S.D.) ∆F/F0 of 1.81 ± 0.30. in chow fed to 1.60 ± 0.32 in LVAD fed mice, respectively (Mann-Whitney test p < 0.05. Supplementary Table S3). Similarly, the peak calcium transients induced by tetanic stimulation of soleus fibers from LVAD mice was decreased from a mean (± S.D.) ∆F/F0 of 1.29 ± 0.42 in chow fed mice to 1.10 ± 0.32 in LVAD (Mann-Whitney test p < 0.05) (Figure 3b and Supplementary Table S3).
Supplementary Table 3: Electrical stimulation increases oxidation of proteins in soleus muscles and this effect is exacerbated by the LVAD. View Supplementary Table 3
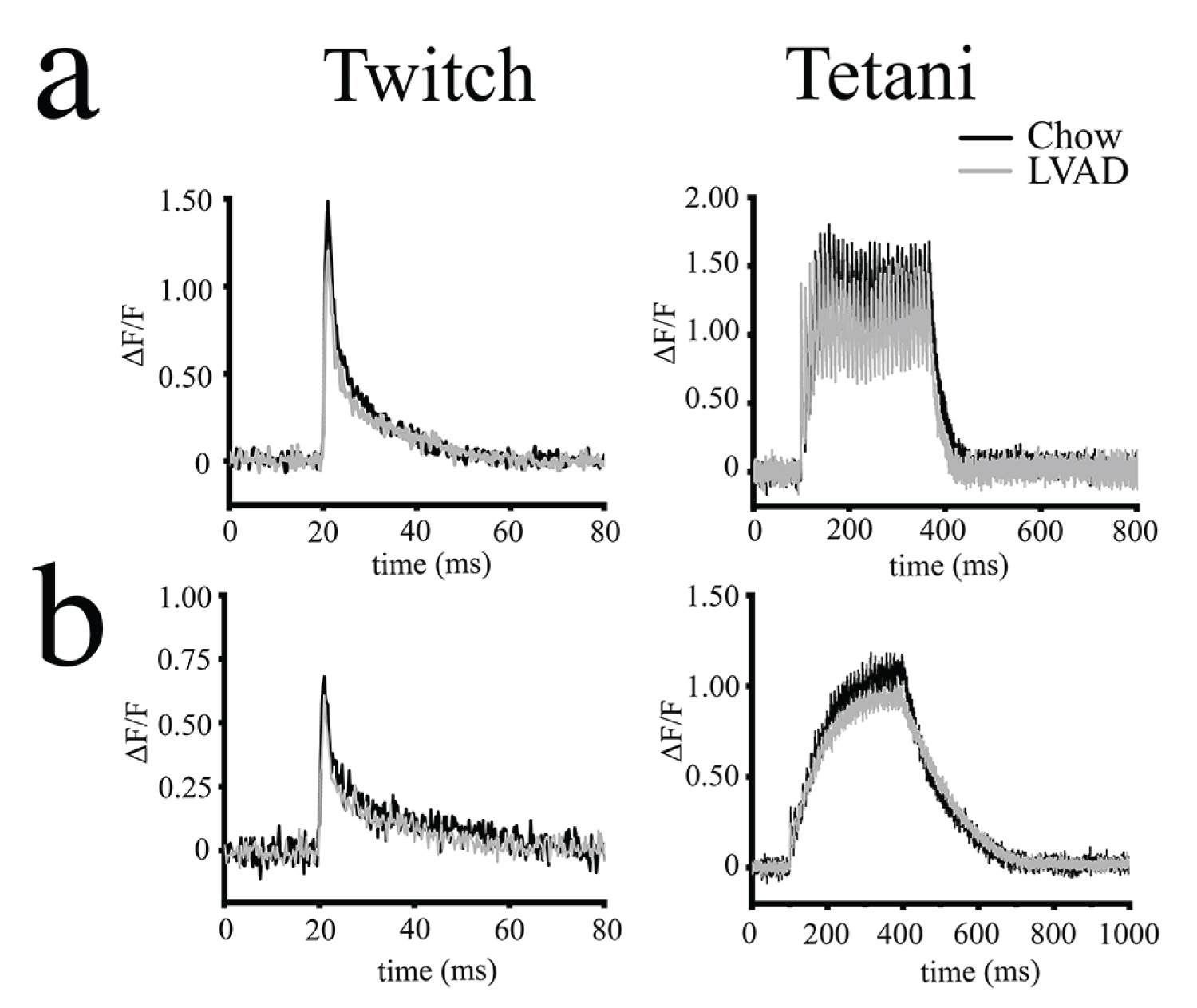 Figure 3: Electrically evoked peak Ca2+ transients are reduced in EDL muscles from LVAD fed mice. Enzymatically dissociated fibres were loaded with Mag-Fluo-4 and electrically stimulated by field stimulation; (a) EDL and (b) Soleus muscles. Left panels show representative Ca2+ transient elicited by a single pulse of 50 V, 1.0 ms duration in. Right panels show representative Ca2+ transient elicited by a train of pulses delivered at 100 Hz for 300 ms. Black trace, chow fed, grey trace, LVAD fed mice.
View Figure 3
Figure 3: Electrically evoked peak Ca2+ transients are reduced in EDL muscles from LVAD fed mice. Enzymatically dissociated fibres were loaded with Mag-Fluo-4 and electrically stimulated by field stimulation; (a) EDL and (b) Soleus muscles. Left panels show representative Ca2+ transient elicited by a single pulse of 50 V, 1.0 ms duration in. Right panels show representative Ca2+ transient elicited by a train of pulses delivered at 100 Hz for 300 ms. Black trace, chow fed, grey trace, LVAD fed mice.
View Figure 3
Vitamin A together with vitamin C and vitamin E have anti-oxidant activity and enhanced oxidative stress has been postulated as being responsible for aging, disease as well as decreased muscle function [15-17]. Since in vivo and ex vivo muscle function is negatively impacted by the LVAD, we investigated whether this is paralleled by enhanced skeletal muscle protein oxidation. Oxidized proteins can be visualized by immunoblotting using the Oxyblot detection system, which derivatives carbonyl groups formed during protein oxidation, to protein hydrazones by incubating them with 2,4-dinitro-phenylhydrazine (DNPH) [18]. Muscles of sedentary mice were isolated, total homogenates were prepared, proteins were derivatized with DNPH and immediately flash frozen in liquid nitrogen. Subsequently, oxidation of proteins was evaluated by western blotting using anti-DNPH antibodies. Figure 4a shows representative Oxyblots (left panels) and densitometric scans (right panels) of the immunopositive bands present in total homogenates from EDL and soleus muscles isolated from chow and LVAD fed sedentary mice. The lower panels show MyHC loading controls. Both EDL and soleus muscles from chow and LVAD fed mice contain a number of oxidized proteins having a wide range of molecular weights. There was no difference in the proportion of oxidized proteins between the chow and LVAD mice (Figure 4a). The high molecular weight oxidized protein present in the total homogenates likely corresponds to MyHC isoforms and their modification would impact force generation. To this aim we separated the MyHC isoforms by running high resolution acrylamide gels [13], blotted them onto nitrocellulose and probed the membranes with anti-DNPH antibodies; after visualization of the immunopositive bands the blots were stripped and re-probed with an anti-MyHC antibody recognizing all isoforms. There was no difference in the level of oxidation of MyHC II b (EDL) or MyHCI and IIa (soleus) between chow and LVAD fed mice (Supplementary Figure S3a).
 Figure 4: Electrical stimulation causes an increase in oxidized protein content in soleus muscles from LVAD fed mice. (a) Muscle homogenates were treated with DNPH and flash frozen. Ten µg total homogenate protein were separated on a 10% SDS PAGE, blotted onto nitrocellulose and probed with the anti-DNPH antibody as described in the Methods section. Positive proteins were visualized by chemiluminescence. The blot was subsequently stripped and re-probed with anti-MyHC (all) for normalization (bottom). Left panels, representative blots; right panel, densitometric scan of the blot; (b) Muscles from chow and LVAD fed mice were isolated and stimulated in vitro by field stimulation (150 Hz for EDLs and 120 Hz for soleus muscles for 10 min). Total homogenates were prepared, treated with DNPH and flash frozen. Ten micrograms of protein were separated on a 10% SDS PAGE, blotted onto nitrocellulose and probed with the anti-DNPH antibody. Positive proteins were visualized by chemiluminescence. The blot was subsequently stripped and re-probed with anti-MyHC (all) for normalization. Left panels, representative blots; right panel, densitometric scan of the blot. The blots shown in panels (a) and (b) are from the same gels but were cropped in order to facilitate comparison of Chow and LVAD, EDL and soleus muscle immunoreactivity. Experiments were performed on muscles from 3 chow fed and 3 LVAD fed mice with similar results.
View Figure 4
Figure 4: Electrical stimulation causes an increase in oxidized protein content in soleus muscles from LVAD fed mice. (a) Muscle homogenates were treated with DNPH and flash frozen. Ten µg total homogenate protein were separated on a 10% SDS PAGE, blotted onto nitrocellulose and probed with the anti-DNPH antibody as described in the Methods section. Positive proteins were visualized by chemiluminescence. The blot was subsequently stripped and re-probed with anti-MyHC (all) for normalization (bottom). Left panels, representative blots; right panel, densitometric scan of the blot; (b) Muscles from chow and LVAD fed mice were isolated and stimulated in vitro by field stimulation (150 Hz for EDLs and 120 Hz for soleus muscles for 10 min). Total homogenates were prepared, treated with DNPH and flash frozen. Ten micrograms of protein were separated on a 10% SDS PAGE, blotted onto nitrocellulose and probed with the anti-DNPH antibody. Positive proteins were visualized by chemiluminescence. The blot was subsequently stripped and re-probed with anti-MyHC (all) for normalization. Left panels, representative blots; right panel, densitometric scan of the blot. The blots shown in panels (a) and (b) are from the same gels but were cropped in order to facilitate comparison of Chow and LVAD, EDL and soleus muscle immunoreactivity. Experiments were performed on muscles from 3 chow fed and 3 LVAD fed mice with similar results.
View Figure 4
 Supplementary Figure 3: Oxyblots of high resolution SDS PAGE of MyHC in EDLs and soleus muscles. (a) Resting muscles: No difference in MyHC IIb oxidation in EDL muscles nor of MyHC I and IIa in soleus muscles was apparent between chow and LVAD fed mice. (b) Electrically stimulated muscles. Electrical stimulation resulted in a significant increase of the level of oxidized MyHC IIB (EDL muscles) and MyHC I and IIa (soleus muscles) in LVAD fed mice. Top blots, anti-DNPH bottom blots anti-MyHC all. Derivatization control solution (DCS) is the negative control in which oxidized proteins are not reacted with DNPH. For the electrical stimulation experiments, EDL and soleus muscles from chow and LVAD fed mice were isolated and stimulated in vitro by field stimulation (150 Hz for EDLs and 120 Hz for soleus muscles for 10 min at a time between stimulus frequency of 0.27 Hz). The bar graphs (right of the western blot) show the DNPH levels normalized by MyHC per MyHC IIb (EDL) and MyHC I and IIa (soleus). * p < 0.05 Mann-Whitney test.
View Supplementary Figure 3
Supplementary Figure 3: Oxyblots of high resolution SDS PAGE of MyHC in EDLs and soleus muscles. (a) Resting muscles: No difference in MyHC IIb oxidation in EDL muscles nor of MyHC I and IIa in soleus muscles was apparent between chow and LVAD fed mice. (b) Electrically stimulated muscles. Electrical stimulation resulted in a significant increase of the level of oxidized MyHC IIB (EDL muscles) and MyHC I and IIa (soleus muscles) in LVAD fed mice. Top blots, anti-DNPH bottom blots anti-MyHC all. Derivatization control solution (DCS) is the negative control in which oxidized proteins are not reacted with DNPH. For the electrical stimulation experiments, EDL and soleus muscles from chow and LVAD fed mice were isolated and stimulated in vitro by field stimulation (150 Hz for EDLs and 120 Hz for soleus muscles for 10 min at a time between stimulus frequency of 0.27 Hz). The bar graphs (right of the western blot) show the DNPH levels normalized by MyHC per MyHC IIb (EDL) and MyHC I and IIa (soleus). * p < 0.05 Mann-Whitney test.
View Supplementary Figure 3
We also investigated whether muscle use impacts the oxidation status of proteins by subjecting isolated muscles to ex vivo electrical stimulation. Electrical stimulation didn't affect the overall content of oxidized proteins in EDL muscles from chow and LVAD fed mice (Figure 4b and Supplementary Table S4). However, Oxyblot analysis showed increased oxidation of MyHC IIB in LVAD mice following electrical stimulation (Supplementary Figure 3b). On the other hand, ex vivo electrical stimulation resulted in a higher proportion of oxidized proteins in soleus muscles from LVAD mice compared to chow fed mice (Figure 4b and Supplementary Table S4). In addition, oxidation of MyHC I and MyHC IIa was significantly increased in LVAD fed mice after electrical stimulation (Supplementary Figure 3b). In order to have a more quantitative view we compared the relative content of oxidized proteins in EDL and soleus muscles before and after electrical stimulation. EDL muscles did not show significant variations in their content of oxidized proteins whereas electrical stimulation significantly increased the relative content of oxidized proteins in soleus muscles from LVAD fed mice (Supplementary Table S4).
Supplementary Table 4: List of antibodies used for western blotting. View Supplementary Table 4
Fatigue is defined as the decline in ability of a muscle to generate force [19] and although its molecular basis is complex, it is thought to be caused by (i) A decrease in metabolic fuel, (ii) The accumulation of metabolites, (iii) A combination of the two plus additional factors [19,20]. Fast twitch muscles are classified as "fatigable" whereas slow twitch muscles are fatigue-resistant [21]. Muscle fatigue can be assessed experimentally ex vivo by measuring the percentage of force developed by isolated muscles after continuous stimulation by a train of action potentials, compared to the force developed in response to the initial stimulation [22]. We next investigated if the LVAD affects muscle fatigue development. Figure 5a shows a representative trace of the force developed by EDL (top traces) and soleus (bottom traces) muscles isolated from chow fed (continuous line) and LVAD fed (dashed line) mice, at the beginning of the stimulation protocol (left trace), after the 20th (EDL), 60th (soleus) tetanic stimulation (middle trace) and after the 70th (EDL), 100th (soleus) tetanic stimulation (right trace). Figure 5b shows the mean (± S.D.) normalized force developed by isolated muscles as a percentage of the initial force developed by the muscle, versus the number of tetani. EDLs were equally sensitive to fatigue in that the force dropped to 10 ± 3% of the initial force after 50 tetani in chow and LVAD fed mice. On the other hand, soleus muscles from LVAD fed mice were more fatigable than those isolated from chow fed mice and the difference was significant after the 70th tetanus. The % normalized force dropped to 30 ± 8% of the initial force after 100 tetani in LVAD fed mice versus 53 ± 7% in chow fed mice, respectively (Figure 5b, right panel).
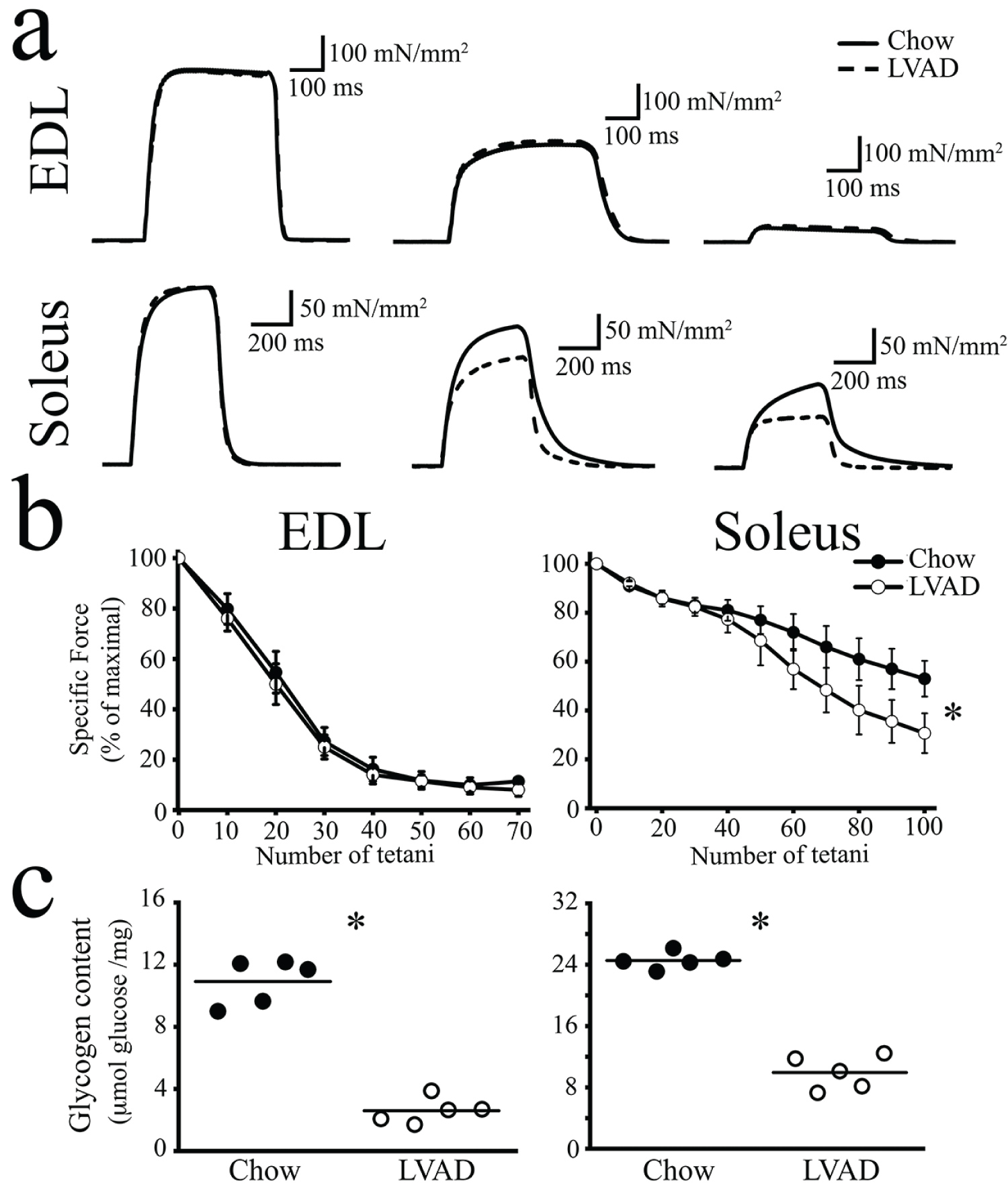 Figure 5: Soleus muscles from LVAD fed mice fatigue earlier and have smaller glycogen stores compared to muscles isolated from chow fed mice. (a) Representative traces of electrically evoked force developed by isolated EDL and soleus muscles with a train of tetani (150 Hz, 300 ms, for 5 min and 120 Hz, 300 ms during 8 min for EDL and soleus, respectively) delivered at 0.27 Hz. Continuous line, chow fed mice; dashed line, LVAD fed mice. Traces represent force developed after the 1st, 20th and 70th tetanic stimulation in EDLs and 1st, 60th and 100th tetanic stimulation in soleus; (b) Normalized mean force versus number of tetani in EDL and soleus muscles. EDLs from chow and LVAD fed mice showed similar fatigue sensitivity whereas soleus muscles from LVAD fed mice were more susceptible to fatigue. Each point represents the mean ± S.D.; n = 5 chow fed and 5 LVAD fed mice; *p < 0.05 Mann-Whitney; (c) Glycogen content was assessed enzymatically in total homogenates from EDL and soleus muscles. Each symbol represents the mean value of both muscles of a single mouse; the median value is represented by the horizontal black line. *p < 0.05 Mann Whitney test.
View Figure 5
Figure 5: Soleus muscles from LVAD fed mice fatigue earlier and have smaller glycogen stores compared to muscles isolated from chow fed mice. (a) Representative traces of electrically evoked force developed by isolated EDL and soleus muscles with a train of tetani (150 Hz, 300 ms, for 5 min and 120 Hz, 300 ms during 8 min for EDL and soleus, respectively) delivered at 0.27 Hz. Continuous line, chow fed mice; dashed line, LVAD fed mice. Traces represent force developed after the 1st, 20th and 70th tetanic stimulation in EDLs and 1st, 60th and 100th tetanic stimulation in soleus; (b) Normalized mean force versus number of tetani in EDL and soleus muscles. EDLs from chow and LVAD fed mice showed similar fatigue sensitivity whereas soleus muscles from LVAD fed mice were more susceptible to fatigue. Each point represents the mean ± S.D.; n = 5 chow fed and 5 LVAD fed mice; *p < 0.05 Mann-Whitney; (c) Glycogen content was assessed enzymatically in total homogenates from EDL and soleus muscles. Each symbol represents the mean value of both muscles of a single mouse; the median value is represented by the horizontal black line. *p < 0.05 Mann Whitney test.
View Figure 5
Since fatigue may be caused by the lack of metabolic fuel availability and we previously reported that SRP-35 over-expression enhances glucose uptake [5], we also assessed muscle glycogen store content. The LVAD significantly affected the total glycogen content of both EDL and soleus muscles (Figure 5c). In particular, EDLs from chow fed mice contained 11.0 ± 1.4 µmoles glucose/mg muscle whereas those from LVAD fed mice contained approximately one quarter, that is 2.5 ± 0.8 µmoles glucose/mg muscle. Soleus muscles have larger glycogen stores and indeed soleus muscles from chow fed mice contained 24.5 ± 1.8 µmoles glucose/mg muscle whereas those from LVAD fed mice contained less than half, that is 9.9 ± 2.2 µmoles glucose/mg muscle.
The results of this study demonstrate that a chronically low vitamin A diet significantly impacts in vivo and ex vivo muscle function. This is likely mediated by a combination of events including increased oxidation of proteins in soleus muscles during activity and decreased glycogen stores, with little or no effect on the content of proteins involved in skeletal muscle ECC. This study supports and extends our previous findings on the role of vitamin A in muscle function, where we showed that producing more retinoic acid locally leads to increased glucose uptake resulting in larger glycogen stores [5].
Vitamin A deficiency negatively affects a number of physiological parameters including eye sight, immune reactions and growth, but little is known regarding its impact on skeletal muscle function. An in-depth study on vitamin A deficiency showed that rats grew more slowly and their body weight was lower when they were fed a vitamin A deficient diet even though food consumption was equivalent to that of chow fed rats. The study concluded that the effect was likely due to muscle catabolism [3]. The LVAD used in this study contained vitamin A albeit, reduced amounts (4 IU/g instead of 20 IU Vitamin A/g) and mice were maintained on this low vitamin A diet to induce a decrease in stored retinoic acid in various tissues [9]. The LVAD did not result in alterations of muscle weight, nevertheless it did impact muscle function. Indeed, LVAD fed mice ran less than chow fed mice and their muscles developed less force when subjected to electrical stimulation. The reduced in vivo muscle function may by caused at least in part by the smaller glycogen stores present in muscles from the LVAD fed mice but this is unlikely to be the cause of the reduced force developed after electrical stimulation nor to the decrease in the peak calcium transient induced by twitch and tetanic stimulation. The LVAD did not cause major changes in the content of RyR1, Cav1.1 or calsequestrin, that is of the major proteins involved in skeletal muscle ECC [23], indicating that the effect of LVAD on force and calcium homeostasis must have additional causes. In this context it should be pointed out that muscles from LVAD fed mice exhibited a decreased content of the calcium buffering protein parvalbumin in fast twitch muscles and an increase of SERCA1 in slow twitch muscles. The reduced parvalbumin content would reduce the speed of relaxation [24,25], paradoxically leading to an increase in the force generated after tetanic stimulation, which was in fact, not observed most likely because the change in parvalbumin content was not large enough [24]. On the other hand, the increase in SERCA1 content in slow twitch muscles could potentially indicate an incomplete switch to a slow fiber type isoform. However, we did not observe changes in MyHC isoform content an event which would support the presence of fast twitch fibers within soleus muscles. Additionally, the increased SERCA1 content was not paralleled by faster kinetics of the calcium transient nor of muscle relaxation, indicating that the changes in the content of these two proteins cannot account for the observed functional ECC changes.
Since the mechanical properties of isolated muscles were affected by the LVAD, but were not influenced by over-expression of SRP-35 [5], one can assume that the LVAD diet must impact muscle function by a different mechanism. To this end we focused on the anti-oxidant function of vitamin A [1,2]. Increased oxidation and oxidative stress have been reported to affect muscle function in different ways, in particular: (i) Supplementation of rodent chow with retinyl palmitate in combination with exercise training adversely affects muscle function by increasing skeletal muscle oxidative stress and decreasing exercise-induced induction of endogenous antioxidant enzymes [26]; (ii) Exposure of muscle fibers to exogenous oxidants such as H2O2 and DTDP causes an increase in Ca2+ sensitivity of the contractile apparatus and contributes to the effect of post-tetanic potentiation [27]; (iii) Reactive oxygen species (ROS) play a role in muscle fatigue [28]. Keeping these facts in mind we analyzed the effects of the LVAD on both the oxidation status of muscle proteins as well as the fatigability of muscles. Oxyblot analysis revealed that even at rest EDL and soleus muscles contain a proportion of oxidized proteins and that the overall extent of protein oxidation in EDL muscles in chow and LVAD fed mice is unchanged even after electrical stimulation. Nevertheless, the extent of MyHC oxidation following electrical stimulation was significantly higher in LVAD than chow fed mice. Interestingly, electrical stimulation didn't increase the content of oxidized proteins in soleus muscles from chow fed mice, though soleus muscles from LVAD fed mice consistently contain a higher proportion of oxidized proteins including a higher proportion of oxidized MyHC I and IIa. These results indicate that under normal conditions (i) The presence of oxidized proteins doesn't necessarily negatively impact muscle function and (ii) That skeletal muscles are equipped with sufficient amounts of anti-oxidants to counterbalance potential excessive protein oxidation resulting from increased muscle use.
The results of the present investigation also raise an interesting question, that is why are the force and calcium transients impacted to a greater extent in fast twitch muscles than in slow twitch muscles in LVAD mice even though slow twitch muscles exhibit a higher content of oxidized proteins? Since the content of ECC proteins in EDL or soleus muscles is not affected by the LVAD we postulate that additional factors are responsible for the decreased muscle function. One such possibility is that the RyR1 and or Cav1.1 whose relative content is higher in fast twitch muscles than slow twitch muscles [10,29] may be subjected to post-translational modifications in LVAD fed mice, leading to a less efficient ECC and to less calcium being release after electrical stimulation, leading to reduced muscle contracture.
In conclusion our results show that vitamin A plays an important role in skeletal muscle function where it has a dual role, both as an anti-oxidant as well as a regulator of glucose uptake and glycogen storage.
This work was supported by funds from Swiss Muscle foundation.
A.R., C.B., M.F., S.B., performed the experiments and analyzed the data; A.R, F.Z. and S.T., drafted the article; A.R., C.B., M.F., S.B., F.Z and S.T., critically revised the manuscript; F.Z. and S.T., supervised all the steps of the study.
The authors declare no competing financial interests.