The hypothesis that early degeneration of the pedunculopontine nucleus (PPN) may be related to nigral degeneration has opened new avenues of basic research for this disease. It is not known how the motor function and nigrostriatal gene expression of TH, VMAT2 and DAT behave in an intermediate term (30 days) following the pontine lesion.
The effect of unilateral PPN neurotoxic injury on forelimb use and basic walking pattern was studied. Likewise, the effect of the pontine lesion on the nigrostriatal gene expression of TH, VMAT2 and DAT was addressed. Both behavioral and molecular studies were performed 30 days after PPN injury.
Behavioral study revealed the motor disabilities associated with the vertical exploration and the gait pattern after 30 days of performing the unilateral neurotoxic lesion of the PPN. The PPN lesioned rats showed a percentage of use the forelimb ipsilateral to the lesion and use of both forelimbs simultaneously, significantly greater than control groups (p ≤ 0.001 and p ≤ 0.05 respectively). The percentage of use of the contralateral forelimb revealed a tendency to decrease in the lesioned group. In connection with basic walking pattern, the stride length, intrastep distance and the gate base of support were significantly decreased (p ≤ 0.05) in the lesioned rats. The spreading toes (1-5) for both, the right (p ≤ 0.05) and left (p ≤ 0.01) limbs showed a significant increase in this group. The spreading inners toes (2-4) for both, the right (p ≤ 0.05) and left (p ≤ 0.05) limbs were also significant increase in lesioned rats. The molecular study revealed a significantly increase in TH (p ≤ 0.05) and DAT (p ≤ 0.01) mRNA nigrostriatal expression together a decrease in VMAT 2 (p ≤ 0.05) gene expression.
Motor dysfunctions associated with the pontine lesion seem to be already irreversible 30 days after the lesion. Reduced VMAT2 gene expression may be an early damage signal that could induces other changes in dopaminergic signaling, 30 days after pontine injury.
Pedunculopontine nucleus, Motor asymmetry, Basic walking pattern, Tyrosine hydroxylase, Vesicular dopamine transporter, Dopamine transporter
PD: Parkinson's Disease; PPN: Pedunculopontine Nucleus; SNpc: Substantia Nigra pars compacta; TH: Tyrosine Hydroxylase; VMAT2: Vesicular Dopamine Transporter; DAT: Dopamine Transporter; CT: Cylinder Test
During the first 30 years after the dopaminergic hypothesis for Parkinson's disease (PD) was proposed, most of the basic research was directed at nigral dopaminergic neurons, since their degeneration constitutes the distinctive neuropathological feature of the disease [1]. It is well known that dopaminergic neurons present multiple risk factors that make them prone to neurodegeneration [2-4]. Diverse experimental models of PD (neurotoxic and genetic) most of them focused on the death of nigral dopaminergic neurons were developed [5,6]. Although these models have been very useful for reproducing some signs and symptoms of the disease, one of their main limitations is that they do not reproduce the slow and progressive course of dopaminergic degeneration [5].
Around 1992, the literature indicated that the excite toxic lesion of the pedunculopontine nucleus (PPN) could generate better experimental models of PD than the standardized models up to that time [7]. Later, work of Braak demonstrated the presence of Lewy bodies in the PPN before nigral degeneration was observed [8]. Based on this knowledge, the number of studies addressing the morphofunctional relationship between the substantia nigra pars compacta (SNpc) and the PPN has grown exponentially [9-11].
The PPN and the SNpc share physiological properties that suggest that both neuron populations are mutually dependent for their survival [10]. The PPN consists of glutamatergic, cholinergic, and GABAergic cells [12,13]. Cholinergic-glutamatergic pontine-nigral output has been attributed a modulatory role of nigrostriatal dopaminergic activity [11].
As background to this study, our group has previously published that the unilateral neurotoxic lesion of the PPN is accompanied by early disorders of gait and motor coordination [14,15]. Similarly, pontine injury is associated with compromised nigrostriatal redox homeostasis that can accentuate nigral vulnerability [14,15]. Also at the molecular level, our previous findings demonstrate changes in the gene expression of both transcription factors and genes involved in the phenotype and dopaminergic homeostasis that together suggest an increased risk condition more prone to degeneration [16,17].
However, these studies have been carried out in a very early time window following the neurotoxic injury of the PPN. In general, these results describe the nigrostriatal biochemical and molecular scenario from the first 24, 48 hours to seven days after the PPN injury. However, it is not known how these changes evolve in a longer time after the neurotoxic injury of the PPN. The goals of the present work, a continuation of what we had previously published, were i) To evaluate the skills in the use of the forelimbs in a vertical exploration task and the changes in the basic walking pattern, ii) The nigrostriatal gene expression of the tyrosine hydroxylase (TH), vesicular dopamine transporter (VMAT2), dopamine transporter (DAT) as well as the ratio DAT/VMAT2, 30 days after pontine injury.
Male Wistar rats weighing 250-300g, from the Center for Breeding of Laboratory Animals (CENPALAB), Mayabeque, Cuba, were housed 5 per cage under a temperature of 22-24 ℃, with a relative humidity of 60% ± 5% and and cycles of 12 hours of light and 12 hours of darkness. Water and food were provided ad libitum. Experiments were carried out in accordance with the Cuban Regulations for the Use of Laboratory Animals (CENPALAB 1997). Research Ethics Committee of International Center of Neurological Restoration (CIREN) analyzed and approved the protocol in February 2021 (Code: ET.7-2021(2)).
The rats were anesthetized by intraperitoneal (i.p) injection a cocktail of ketamine (mg/ml), diazepan (mg/ml) and atropine (mg/ml) in a dose of 1 mL/100g and placed in a frame of stereotactic surgery (David Kopf Instruments, Tujunga, CA, USA). Our group has already published the details of the procedure followed in the surgery [17-19]. In brief: the neurotoxic lesion of the PPN was performed by means of the injection of an N-methyl-D-aspartate (NMDA) (Sigma, St. Louis, MO, USA) solution in the coordinates (mm) corresponding to the PPN, according to the 20- Paxinos and Watson atlas [20] AP: -7.80 ML: 1.60 DV: -7.60. At the end of the surgical procedure, the rats remained in postoperative care until their complete recovery. Sham lesion of the PPN (sham-operated group): The surgical procedure was the same but in place of the neurotoxin the rats received 0.5 μL of physiological saline solution. Figure 1 shows the temporal scheme of the experimental design.
 Figure 1: The experimental design focuses on unilateral neurotoxic injury of the pedunculopontine nucleus. Thirty days after the injury, behavioral studies were performed. First the footprint test and then the cylinder test. Once the behavioral studies were completed, the rats were sacrificed, the tissues were dissected and the molecular biology study was carried out.
View Figure 1
Figure 1: The experimental design focuses on unilateral neurotoxic injury of the pedunculopontine nucleus. Thirty days after the injury, behavioral studies were performed. First the footprint test and then the cylinder test. Once the behavioral studies were completed, the rats were sacrificed, the tissues were dissected and the molecular biology study was carried out.
View Figure 1
Cylinder test: The cylinder test (CT) allowed the evaluation of asymmetry in the use of the forelimbs during a vertical exploration task. The rats were individually placed inside a transparent cylinder (20 cm in diameter and 30 cm high). The behavioral testing was performed during the dark portion of the light cycle. The number of contacts with the cylinder wall made with the forelimb ipsi and contralateral to the lesion separately as well as the number of contacts made with both extremities simultaneously were recorded. At the same time, latency to first contact was registered, as the time it takes for the rat to make first contact with any of its forelimbs on the cylinder wall. The percentage of use of the (right, left and both) forelimb relative to the total contact was calculated. The ratio of contacts (index right asymmetry) made by the ipsilateral to the lesion forelimb relative to the total number of contacts (ipsilateral + contralateral + both forelimbs simultaneously) was calculated using the formula: [(ipsilateral limb contacts + 0.5 both limb contacts)/total number of contacts]. Scores above 0.5 indicate a greater reliance in that limb during the execution of the vertical exploration task [21].
Foot print test: Footprint analysis was performed to evaluate the changes in the basic walking pattern. Rats were manually restrained and both the hind paws were immersed in nontoxic ink. The rats were then allowed to walk freely in a restricted straight corridor (diameter: 17 cm; height: 25 cm). The floor of the gangway was lined with conventional paper, and the footprints marked on the paper were analyzed. Two groups of variables were analyzed. Stride length (mean distance between consecutive right and left paw prints), gate base of support (mean distance perpendicular to parallel left and right paw prints), intrastep distance (mean distance between alternate left and right paw prints) facilitates the study of the basic walking pattern globally. On the other hand, spreading inners toes (toes 2-4), spreading toes (toes 1-5) and foot length (area touched from toe 3 to other end of the foot) make possible to evaluate the position adopted by the toes and hindlimb itself during locomotion [15,22].
Sample collection: Our group has already published the details of this procedure previously [17,18]. Briefly: the rats were anesthetized by intraperitoneal (i.p) injection a cocktail of ketamine (mg/ml), diazepan (mg/ml) and atropine (mg/ml) in a dose of 2 mL/100 g. The animals were sacrificed by decapitation. The cranial bones and meninges were carefully removed. The brains were washed in physiological saline solution and placed on an ice-cooled glass plate. The dissections of the tissues were carried out under stereoscopic microscope (Wild Heerbrugg Stereo Microscopes, Heerbrugg, Switzerland). The molecular studies were performed on the right nigrostriatal tissue, ipsilateral to the PPN lesion. All of the brain tissues were stored at -80 ℃ until analysis.
Transcription factors mRNA expression was studied at 30 days after the PPN neurotoxic lesion. The total RNA was extracted from the nigral and striatal tissue of 15 rats using TRIzol® Reagent (Invitrogen, Carlsbad, CA, USA). The samples were eluted in 30 μL de Ultra Pure TM Distiled Water (Invitrogen, Waltham, Massachusetts, USA) and quantified in the in the Nano Drop One Cspectrophotometer (THERMO Scientific, Waltham, Massachusetts, USA) at ƛ = 260 nm. Samples had 260-280 ratios in the range 1.8-2.1 they were selected, and their integrity was determined by 1% agarose gel electrophoresis. The RNA concentrations were registered, and the samples were stored at -70 ℃. To synthesize cDNA, 1 μg of total RNA was reverse transcribed with ImProm-IITM Reverse Transcription System following the manufacturing protocol without modifications (PROMEGA, USA). The PCR was performed at Select Cycler II (Select BioProducts, Wembley, Western Australia, Australia) using 3 μL of cDNA mixed with 2 mM dNTPs, 6 mM MgCl2, 0.4 μM for each primer and 1U of Taq DNA polymerase (RadiantTM, Alkali Scientific Inc, Fort Lauderdale, Florida, USA). Cycling conditions were as follows: 95 ℃ for 1 min; 40 cycles of 95 ℃ for 15 s; 64 ℃ (annealing temperature) for 1 min; 72 ℃ for 1 min; one last step of 72 ℃ for 10 min. The sequence of each primer (forward and reverse primers sequences (5´ to 3´) and the length of the amplified products were as follows: TH forward: TTC CCC ATG TTC AAC GGA CC, reverse: GCG AGC ACA GTA ATC ACC TTC, product length (bp): 449. VMAT2, forward: CGC AAA CTG ATC CTG TTC AT; reverse: AGA AGA TGC TTT CGC AGG TG, product length (bp): 175. DAT, forward: GGA AGC TGG TCA GCC CCT GCT T; reverse: GAA TTG GCG CAC CTC CCC TCT G, product length (bp): 266. β-Actin was employed as endogenous control, forward: ATT TGG CAC CAC ACT TTC TAC A; reverse: TCA CGC ACG ATT TCC CTC TCA G, product length (bp): 379. PCR products were separated by 2% agarose/ethidium bromide gel electrophoresis and visualized under UV light. Each electrophoresis was performed twice, followed by a semi quantification analysis. For the semi-quantitative study, the free online program ImageJ (Version1.44; Wayne Rasband, National Institute of Health, Bethesda, MD, USA; http://imagej.nih.gov/ij) was employed. The analysis was performed according to the published method [23]. The background activity of the target band was subtracted and then normalized using β-actin as a reference.
Normal distribution and homogeneity of variance of the data were analyzed applying the Kolmogorov-Smirnov and Levene tests, respectively. The comparison between experimental groups of both the variables associated with the behavioral studies (foot print test and cylinder test) and the molecular biology studies (TH, VMAT2 and DAT mRNA expression) was carried out by a one-way analysis of variance (ANOVA) followed by a Tukey Test. For these variables, the values are expressed as mean ± SEM. The analysis of the DAT/VMAT2 mRNA expression ratio was performed by a Kruskal-Wallis test. The behavior of this variable is shown by the median.
For all analyses, a significance level of 0.05 was considered using the Statistica 8.0 software (StatSoft Ink, Tulsa, OK, USA) software. The graphics were prepared using Graph Pad Prism 5.0 software (Graph Pad Software, California, EEUU).
Cylinder test: The cylinder test allowed us to evaluate the asymmetry in the use of the forelimbs during the vertical exploration, 30 days after the pontine lesion. The PPN lesioned rats showed a percentage of use the right forelimb (ipsilateral to the lesion) during the vertical exploration of the cylinder wall significantly greater than control groups (F(2, 34) = 11.35 p ≤ 0.001) (Figure 2A). On the other hand, the percentage of use of the left forelimb during the exploration of the cylinder wall revealed a tendency to decrease in the group of injured rats, although the behavior of this variable did not reach statistical significance (F(2, 34) = 2.40 p > 0.05) (Figure 2A). In connection with the percentage of use of both forelimbs simultaneously, this variable presented a significantly decrease in PPN lesioned rats (F(2, 34) = 4.74 p ≤ 0.05) (Figure 2A).
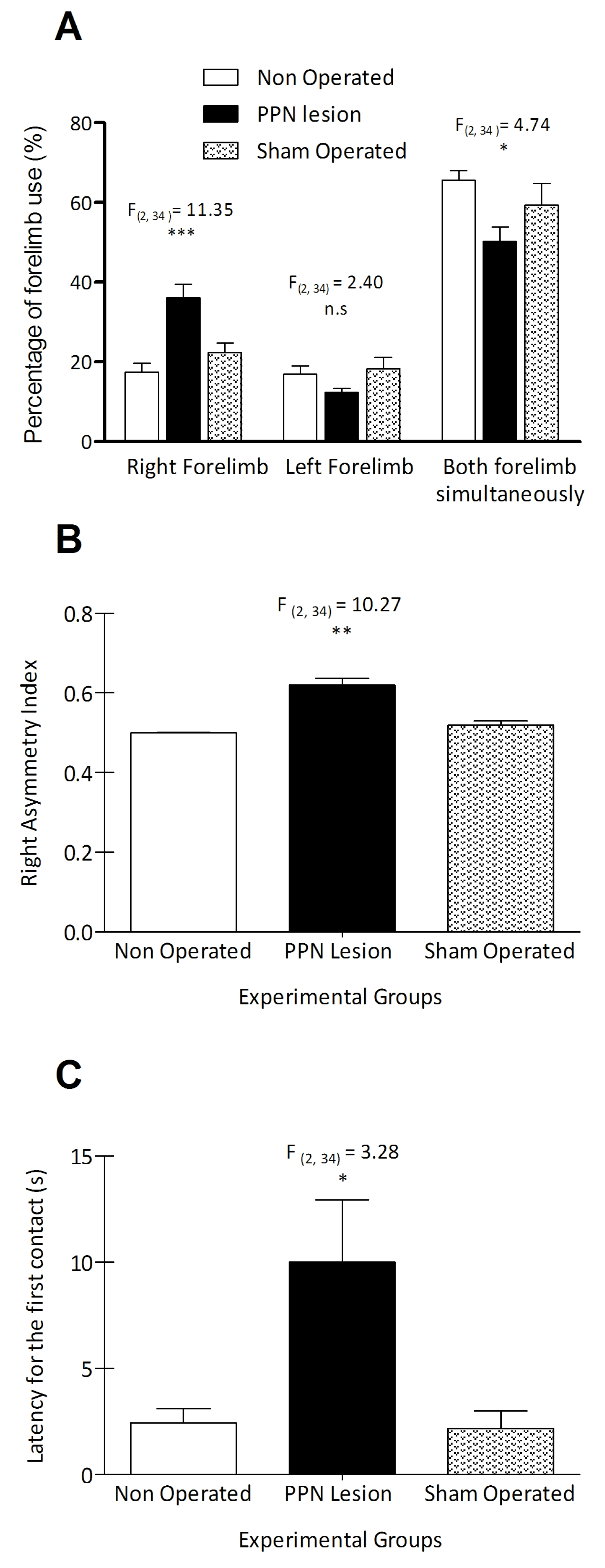 Figure 2: Use of the forelimbs in the cylinder test, 30 days after the unilateral neurotoxic lesion of the PPN. (A), Comparison of the percentage of use of the forelimbs in a vertical exploration task, between experimental groups. (B), Comparison of right asymmetry index between experimental groups. (C), Comparison of the latency to the first contact with the cylinder wall, between experimental groups. Experimental groups: Non-Operated rats (n = 14), PPN lesion (n = 13), Sham-operated rats (n=10). *p ≤ 0.05; **p ≤ 0.01; ***p ≤ 0.001. Bars represent the x ± SEM.
View Figure 2
Figure 2: Use of the forelimbs in the cylinder test, 30 days after the unilateral neurotoxic lesion of the PPN. (A), Comparison of the percentage of use of the forelimbs in a vertical exploration task, between experimental groups. (B), Comparison of right asymmetry index between experimental groups. (C), Comparison of the latency to the first contact with the cylinder wall, between experimental groups. Experimental groups: Non-Operated rats (n = 14), PPN lesion (n = 13), Sham-operated rats (n=10). *p ≤ 0.05; **p ≤ 0.01; ***p ≤ 0.001. Bars represent the x ± SEM.
View Figure 2
Right asymmetry index showed a significant increase for lesioned rats (F(2, 34) = 14.27 p ≤ 0.001) whereas the latency to the first contact with the cylinder wall showed significant difference between experimental groups (F(2, 34) = 3.28 p ≤ 0.05) (Figure 2B and Figure 2C).
Foot print analysis: Thirty days after unilateral PPN neurotoxic injury, rats showed changes in their gait pattern. The analysis of stride length indicated a significant decrease for both the right (F(2, 30) = 5.485 p ≤ 0.05) and the left (F(2, 30) = 3.60 p ≤ 0.05) sides in the lesioned group (Figure 3A). The intrastep distance (F(2, 30) = 3.746 p ≤ 0.05) and the gate base of support (F(2, 30) = 6.60 p ≤ 0.05) also showed a significant decrease in the lesioned rats (Figure 3B and Figure 3C).
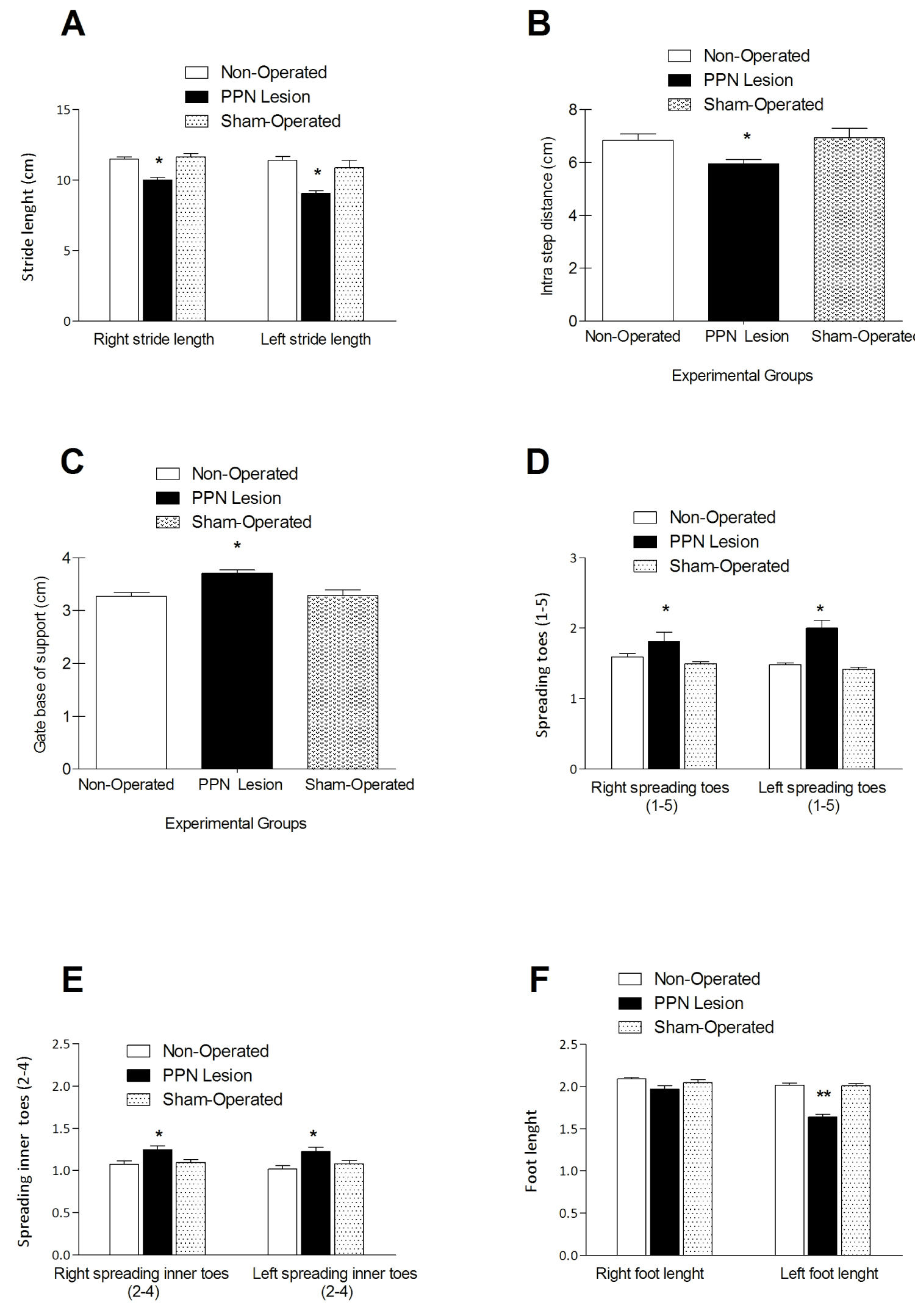 Figure 3: Comparison between experimental groups of the variables that describe the basic walking pattern following 30 days after the unilateral neurotoxic lesion of the PPN. (A), Right and left stride lengths. (B), Intra step distance. (C), Gate base of support. (D), Spreading toes (1- 5). (E), Spreading inner toes (2-4). (F), Foot length. *p ≤ 0.05. Experimental groups: Non-operated (n=10), PPN lesion (n = 10), and sham-operated rats (n = 12). Bars represent mean ± SEM.
View Figure 3
Figure 3: Comparison between experimental groups of the variables that describe the basic walking pattern following 30 days after the unilateral neurotoxic lesion of the PPN. (A), Right and left stride lengths. (B), Intra step distance. (C), Gate base of support. (D), Spreading toes (1- 5). (E), Spreading inner toes (2-4). (F), Foot length. *p ≤ 0.05. Experimental groups: Non-operated (n=10), PPN lesion (n = 10), and sham-operated rats (n = 12). Bars represent mean ± SEM.
View Figure 3
On the other hand the lesioned rats presented a significant increase in spreading toes (1-5) for both, the right (F(2, 30) = 3.735 p ≤ 0.05) and left (F(2, 30) = 8.231 p ≤ 0.01) limbs. This significant increase is maintained for the variable spreading inners toes (2-4) for both, the right (F(2, 30) = 5.663 p ≤ 0.05) and left (F(2, 30) = 8.358 p ≤ 0.05) limbs (Figure 3D and Figure 3E). Finally, the foot length showed a non-significant difference in the right limb (F(2, 30) = 3.09 p > 0.05) and a significant decrease in the left limb (F(2, 30) = 10.16 p ≤ 0.001) (Figure 3F).
Thirty days after pontine lesion the TH mRNA expression was significantly higher than control groups (F(2, 14) = 4.15 p ≤ 0.05) (Figure 4A, Figure 4B, Figure 4C, Figure 4D and Figure 4E). Meanwhile, VMAT2 mRNA expression exhibited a significantly decrease in comparison with control groups (F(2, 14) = 4.15 p ≤ 0.05) (Figure 4 B, Figure 4C, Figure 4D and Figure 4E).
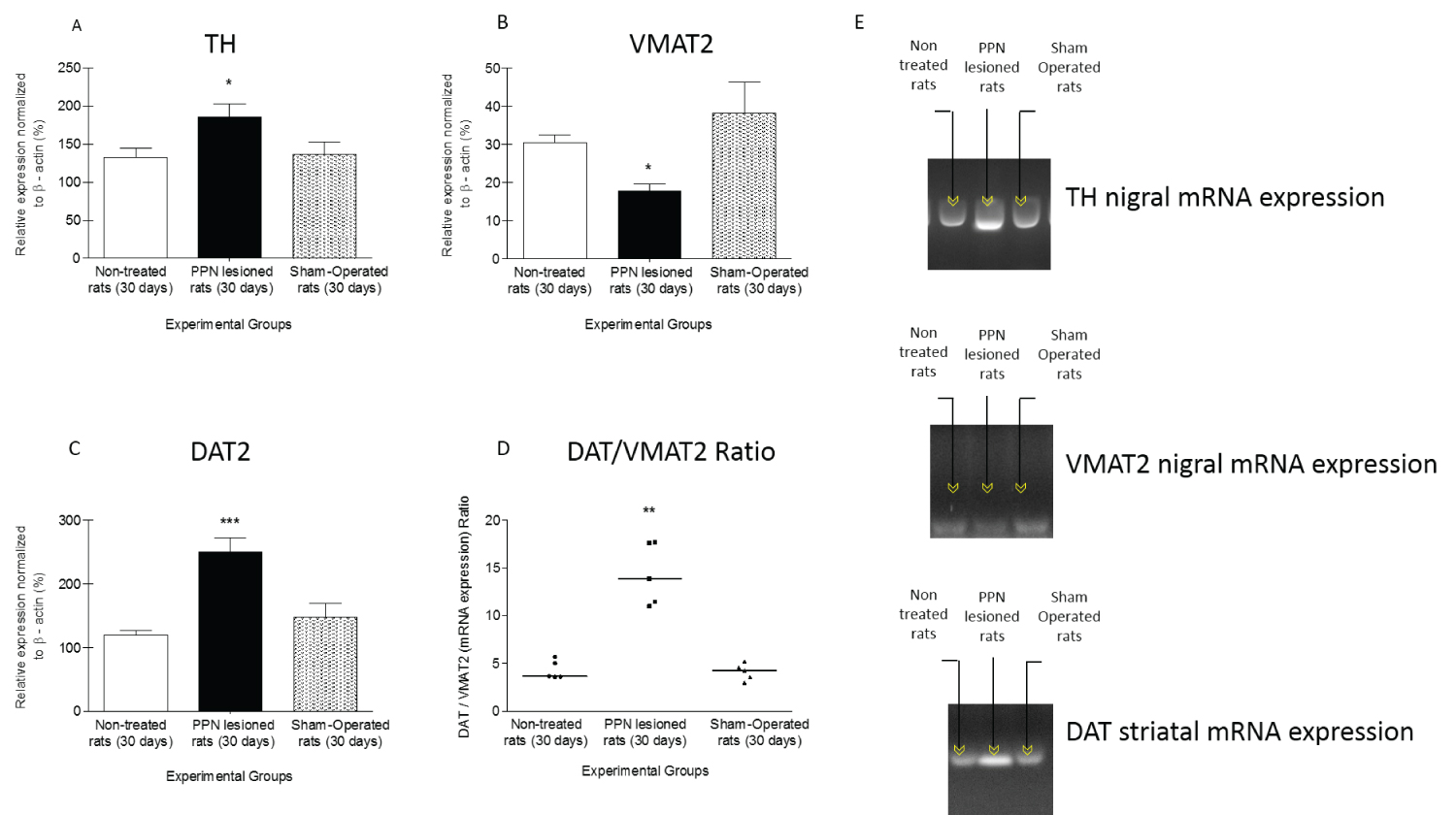 Figure 4: Comparison between experimental groups of nigrostriatal gene expression of proteins related to dopaminergic homeostasis, 30 days after unilateral PPN neurotoxic lesion. The study was by semi-quantitative RT-PCR, expressed as percentage of b-actin. (A) Tyrosine hydroxylase (TH) mRNA expression in nigral tissue. (B) Vesicular monoamine transporter 2 (VMAT2) mRNA expression in nigral tissue. (C) Dopamine transporter (DAT) mRNA expression in striatal tissue. (D) DAT/VMAT2 ratio. (E) Agarose gel electrophoresis bands representative of the semiquantitative RT-PCR study for each protein studied. Experimental groups: Non-operated (n = 5), PPN lesion (n = 5), and sham-operated rats (n = 5). *p ≤ 0.05; **p ≤ 0.01; ***p ≤ 0.001. Bars represent mean ± SEM for A-C. The straight line in panel D represents the median.
View Figure 4
Figure 4: Comparison between experimental groups of nigrostriatal gene expression of proteins related to dopaminergic homeostasis, 30 days after unilateral PPN neurotoxic lesion. The study was by semi-quantitative RT-PCR, expressed as percentage of b-actin. (A) Tyrosine hydroxylase (TH) mRNA expression in nigral tissue. (B) Vesicular monoamine transporter 2 (VMAT2) mRNA expression in nigral tissue. (C) Dopamine transporter (DAT) mRNA expression in striatal tissue. (D) DAT/VMAT2 ratio. (E) Agarose gel electrophoresis bands representative of the semiquantitative RT-PCR study for each protein studied. Experimental groups: Non-operated (n = 5), PPN lesion (n = 5), and sham-operated rats (n = 5). *p ≤ 0.05; **p ≤ 0.01; ***p ≤ 0.001. Bars represent mean ± SEM for A-C. The straight line in panel D represents the median.
View Figure 4
In connection with the DAT mRNA expression, this variable showed a significantly increase in striatal tissue in contrast with control groups (F(2, 14) = 13.23 p ≤ 0.01) Figure 4C, Figure 4D and Figure 4E)).
On the other hand, the DAT/VMAT2 ratio is representative of the dynamic DA uptake and storage process. This variable presented a significant increase 30 days after PPN unilateral neurotoxic lesion in comparison with control groups (H(2, 14) = 9.50 p ≤ 0.01) (Figure 4D).
According our behavioural study the motor disability deepens 30 days after PPN unilateral neurotoxic lesion. The asymmetry in the use of the forelimbs in a vertical exploration task and the changes in the basic walking patter did not show recovery signs after this time post lesion. Likewise, in the temporary window that represents a medium term, we found important changes in the gene expression of TH, VMAT2 and DAT that had not been previously reported. These changes suggest that from a molecular point of view, compromise of pontine nigral cholinergic-glutamatergic innervation can significantly modify nigrostriatal dopaminergic homeostasis.
Medium-term changes in symmetrical forelimb use and basic walking pattern, following 30 days after pontine injury: Cylinder test allow evaluate general aspects of motor control as well as more particular asymmetries. Within the general aspects, we can mention the deterioration in the initiation of the exploratory movement together with the control of the static and dynamic balance, especially during the displacement of the center of gravity. At the same time, the test represents a very useful tool in the evaluation of asymmetry in the use of the forelimbs [24]. Exploration in the cylinder implies a circular movement that requires precise coordination between the support of the forelimbs on the wall of the cylinder and the movement of the hind limbs in a circular direction. The integrity of the propioceptive and motor processing and precise coordination between the fore and hind limbs ensures displacement of the center of gravity of the rat's body in a vertical position, without compromising balance [25].
The results of our study suggest that there is no disuse of the left forelimb (contralateral to the lesion) when comparing the percentage of its use between experimental groups, although this variable shows a descendent trend, without reaching statistical significance. Nevertheless, qualitative movement analysis reveals that injured rats are limited in their ability to correct use the forelimb contralateral to the injury. This is evident in two aspects: i) The position that the limb adopts with the digits flexed against the wall of the cylinder; ii) The paw dragging behavior when the left forelimb drag vertically down the cylinder wall. On the other hand, the injured rats show a significant increase in the percentage of use of the right forelimb (ipsilateral to the lesion) together with a significant decrease in the percentage of use of both limbs simultaneously. Taken together, these results point to a notable asymmetry in the use of the forelimbs that support the value of the right asymmetry index reported in this study.
These results are quite consistent with the results shown by our group for the evaluation of the cylinder test seven days after the unilateral neurotoxic lesion of the PPN [14]. This aspect suggests that the asymmetry in the use of the forelimbs is installed early following the pontine lesion, and is not modified in the medium term (30 days after PPN lesion). On the other hand, the significant increase in latency for the first contact in the lesioned rats group, suggests that 30 days after the pontine damage, the animals develop some hypokinesia. This delay in starting the exploration in a low complexity task, such as the one evaluated by the cylinder test, point to less spontaneous activity in lesioned rats [26].
The correct use of the forelimbs requires proper processing in the neuraxis, which includes the motor cortex and subcortical motor control centers [27]. The nuclei of the reticular formation, including the PPN, play a key role in maintaining postural stability and both static and dynamic balance [27]. The exploration involves movements in a circular direction and movements up and down. In relation to paw dragging behavior, this had already been described in models of focal cortical, ischemic lesion developed in mice [28]. Paw dragging behavior is not observed during circular movements. This behavior is seen only when the digits of the left forelimb are dragged vertically down the cylinder wall. Lesioned rats make asymmetric, preferential use of the limb ipsilateral (right forelimb) to the lesion. However, support of the contralateral limb, although gross, provides postural stability and prevents balance compromise. The unilateral neurotoxic lesion of the PPN early compromises its integrating role in the motor control mechanisms that guarantee muscle tone, balance and postural stability [14]. Thirty days after pontine injury, no compensatory behavioral adjustments suggesting recovery of this integrative function are observed.
Walking is a complex behavior that combines reflex movement patterns and voluntary movements. Its control involves the initiation and termination of locomotion as well as cadence, speed, and coordination between limbs [29]. The footprint test allows the rat's gait pattern and gait cycle to be evaluated [22]. Stride length, gate base of support and intrastep distance make it possible to study the basic walking pattern globally. Meanwhile, spreading inners toes, spreading toes and foot length allow knowing the position adopted by the extremities during locomotion.
Our group published that seven days after the unilateral neurotoxic lesion of the PPN, there are subtle changes in the basic walking pattern of the rats [15]. These changes were characterized by a slight decrease in stride length (right and left), intrastep distance, and support area.
The present work demonstrates that these subtle changes in the variables that characterize the basic walking pattern of the rat, once installed, are not modified 30 days after the pontine injury. The left and right stride length, intrastep distance and gate base of support maintain a significant decrease compared to control rats, 30 days after PPN neurotoxic injury.
Regarding toe position, our group had reported that seven days after PPN injury, no significant differences were found between injured and control rats [15]. However, the significant increase in spreading toes and spreading inner toes together with the decrease in left foot length, 30 days after the PPN injury, represent some of the most interesting findings of the present work.
The basic walking pattern for most mammals, including rodents, consists of the following diagonal limb couplets: right forelimb - left hindlimb - left forelimb - right hindlimb [30]. Morphofunctional studies indicate that the pyramidal tract determine the limb trajectory (forepaw movements and digit use) and guidance [31]. Nevertheless the basic walking pattern is generated from spinal circuits that are under control principally of the reticulospinal tract with certain contribution from the corticospinal and rubrospinal tracts that regulate forelimb placement [31,32]. Meanwhile, rodent hind limbs adapt to changes in forelimb trajectory or center of gravity [31]. The literature suggest the involvement of the PPN in the modulation of the basic gait pattern [33]. Different works indicate that the PPN is part of the ¨mesencephalic locomotor region¨ (MLR) [33,34]. MLR is a functional neuro circuit modulating movement, rhythm, and speed upon electrical stimulation [33,34]. MLR projections modulate activity of motoneurons and the ¨central pattern generators¨ in the spinal cord (via the reticulospinal tract) that regulate the locomotor activity [33]. According to the literature, reticulospinal system would participate in the synergic actions of the neck; trunk and limb muscles while modulate the ¨central pattern generators¨ in the spinal cord [32]. Probably, an asymmetric activation of the reticulospinal tract following neurotoxic PPN injury may contribute to changes in the basic gait pattern.
On the other hand, the differences found in the distance between the inner and outer toes of the injured rats compared to the control groups, adds to the changes in the basic walking pattern. The digits remained open and extended, which could explain the significant increase in the spreading toes and spreading inner toes variables. Toe position during locomotion suggests an adaptive change to counteract the significant decrease in intrastep distance and gate base of support.
Walking and stepping movements are very skillful movements that require the interaction of many nuclei in the brain [35]. The combination of variables that support the forward motion of the body along with the position of the feet and toes ensure fine tuning of the forward motion. This fine-tuning is impaired early in rats with neurotoxic PPN injury and shows no signs of recovery 30 days after injury. The lesion of the PPN can compromise to some extent the multiple connections of this nucleus with structures such as the SNpc, the thalamus and the cerebellum [36,37]. All of them participating in the fine, supraspinal control of the basic walking pattern.
Medium-term changes in nigrostriatal TH, VMAT2 and DAT mRNA expression, following pontine injury: Our finding point to an up regulation of TH and DAT mRNA expression together with a down regulation of VMAT2 in the nigrostriatal system following 30 days after PPN neurotoxic lesion.
TH converts L-tyrosine to L-DOPA as the first and limiting step of cathecolamines synthesis [38]. This enzyme is regulated by complex and multifaceted mechanisms [39]. The literature defines that in the short term, a negative feedback mechanism exerted by dopamine itself participates in the regulation of TH. Phosphorylation and dephosphorylation of the enzyme also participate in the regulation process. Medium to long-term regulation of the enzyme activity involves several stages of gene expression. Under stress conditions, this enzyme increases by induction at the transcriptional level [39-41].
It is very interesting that our group has already published that seven days after PPN lesion there was a discrete increase in TH nigral mRNA expression [18]. Thirty days after pontine lesion this variable maintains the same tendency. Probably, the PPN neurotoxic lesion imposes a tension condition to the pontine-nigral synapse due to the compromise of the glutamatergic and cholinergic components of this projection [10]. It is possible that this condition is accompanied by a functional reorganization of the cholinergic and glutamatergic receptors present at nigral level [42]. Alterations in TH gene expression have been reported in response to the administration of various drugs or manipulation of several endogenous hormone systems [38]. However, the medium-term impact of pontine degeneration on TH nigral gene expression has not been studied. To our knowledge, this is the first work that addresses this aspect.
In this molecular scenario, the increase in nigral TH gene expression cannot be examined as isolated aspect. This change act in concert with the modifications of the VMAT2 and DAT mRNA expression. Together, cause a diminished regulation of dopaminergic homeostasis. Enhanced nigral TH mRNA expression could be source of oxidative stress [43]. This aspect would be potentiated by the higher striatal DAT mRNA expression and lower VMAT2 nigral gene expression in comparison with control groups.
On the other hand, the results of the molecular biology study confirm a significant increase in the DAT/VMAT2 ratio, 30 days after the unilateral neurotoxic lesion of the PPN. This result responds to the severe and prolonged decrease in the nigral VMAT2 gene expression, concomitant with the enhanced striatal DAT gene expression.
VMAT2 and DAT are two regulators of dopamine homeostasis in the presynaptic neuron [44]. VMAT2 is a proton-gradient dependent vesicular transporter meanwhile DAT is a plasma membrane dopamine transporter [45,46]. VMAT2 and DAT perform in concert to control the compartmentalization and movements of dopamine at the synapse [44]. The dynamic and delicate balance established between both transporters, permanently regulate dopamine nigrostriatal neurotransmission [47,48].
Our group has published that seven days after the pontine injury there is a compromise of the nigrostriatal redox homeostasis together with a down regulation of VMAT2 and DAT gene expression [18]. In the present work, our results indicate that the early down regulation of nigral VMAT2 gene expression is already irreversible, 30 days after the pontine injury. However, the up regulation in striatal DAT gene expression could be difficult to explain.
In the cellular and molecular scenario proposed by our work, there is no significant loss of cell bodies in the SNpc. However, our group reported a decrease in the area of the nigral cell bodies, 30 days after the pontine injury, suggestive of an incipient and ongoing degenerative process [17]. In this condition, we are hypothesizing that the lower nigral gene expression of VMAT2 could induce a decrease in striatal DA release. Consequently, different compensatory mechanisms may be manifested, among them, an up regulation of striatal D2 dopamine receptors [49]. The up regulation of D2 leads to activation of ERK and P13K pathway [50]. The activation of these pathways, together, promotes cell surface expression of DAT and therefore enhance dopamine uptake [50,51]. DAT interacts with various proteins, some of which can modulate its expression, membrane distribution and activity [52]. The expression of D2 and DAT in the nigrostriatal axon terminals could represent an anatomical indication of the possible interaction between these two proteins [53]. In turn, an up regulation of DAT mRNA expression could mean a tendency to reconstitute the pool of intracellular dopamine to pack in vesicles and then be released in the synaptic cleft. DAT is responsible for recycling the neurotransmitter back to the presynaptic neuron allowing its reuse. In this role, DAT participates directly to the buildup of cytosolic dopamine and indirectly influences vesicular dopamine as well [46]. However, given the lower nigral gene expression of VMAT2 after pontine lesion, this process may be insufficient. In this case, probably, intracellular DA accumulates, and the risk of cytosolic oxidation and autoxidation increases, deepening the nigral redox compromise [54].
Our results suggest that the changes in the nigrostriatal gene expression of VMAT2 and DAT do not advance in parallel in pontine lesion conditions. Thirty days after the neurotoxic injury of PPN, they cannot still be a temporary window of long term in which all the cellular and molecular changes suggestive of nigral dopaminergic degeneration have fully developed.
The present work is part of the current trends in the search for a model capable of reproducing the slow and progressive course of nigral degeneration. The PPN lesion model proposed in this paper allows the direct and indirect evaluation of the pontine nigral morphofunctional relationship.
Asymmetry in the use of the forelimbs and changes in the basic walking pattern do not change appreciably between 7 and 30 days after pontine injury. Taken together, these findings point to early dysfunction, without changes in the medium term, of the circuits related to the PPN, which connect this structure with other nuclei of the neuraxis. The results of our behavioral studies suggest looking at the relationship of the PPN with the motor cortex, striatum, cerebellum, as well as with the SNpc and other nuclei of the basal ganglia. Each of these connections, separately and all together, including those with a bilateral component, determines the motor asymmetry that encompasses the aspects addressed in this paper.
At the molecular level, our results point directly to changes in nigrostriatal dopaminergic homeostasis, in close association with neurotoxic PPN injury. The behavior of the variables studied suggests that the change in the cholinergic and glutamatergic signalization on the SNpc, resulting from the pontine lesion, depresses the gene expression of VMAT2. In consequence, the vesicular packaging of the DA and its functional availability for the nigrostriatal dopaminergic synapse should be significantly modified. This could constitute a signal that simultaneously increases the gene expression of DAT and TH in an attempt to improve the availability of dopamine either through de novo synthesis or through reuptake from the synaptic cleft. Nevertheless, both converges again in a lower gene expression of VMAT2 that interferes with the normal development of the DA synthesis-vesicular packaging-release-reuptake machinery. A positive feedback loop could deepen and consolidate subsequent damage and, in a longer time window, could reproduce the slow and progressive course of nigral degeneration.
The authors would like to thank the support offered by the CIREN-UAQ Academic Mobility and Collaboration Agreement in which one part of the activities of the present investigation were inserted. At the same time, the authors wish to thank Yamilé Vega for her technical support in the different stages of the experiment.
The authors declare no conflict of interest.
This research did not receive any specific grant from funding agencies in the public, commercial, or not-for-profit sectors.
Conceptualization, L.B. and G.Z.L.L.; methodology, M.E.G.-F.; R.M.P.S. and M.D.F.P; validation, T.S.-S.; L.F.T.; formal analysis, L.B.; investigation, L.B.; M.E.G.-F.; resources, L.B.; data curation, L.B. R.M.P.S.; writing-original draft preparation, L.B; M.E.G.F.; writing-review and editing, L.B.; visualization, L.B.; supervision, L.B.; project administration, L.B.L.; G.Z.L.L.; funding acquisition, L.B. All authors have read and agreed to the published version of the manuscript.