Rapid Eye Movement (REM) sleep deprivation is understood to be associated with more severe negative effects than REM fragmentation. Comparison of the corticosterone response between these patterns of sleep disruption has not been well characterized. Black Swiss mice were exposed to 1-day and 3-day periods of REM deprivation with inverted flower-pot method or REM fragmentation using the moving bar method. Immediately after experimental procedures mice were sacrificed and blood collected. Corticosterone levels were measured in the plasma portion using enzyme-linked immunoassay. Corticosterone levels in both REM deprivation and REM fragmentation were elevated in comparison to baseline, but were not significantly different from each other. In continuous conditions of sleep disruption- either REM deprivation or REM fragmentation- markers of stress are elevated to a similar degree.
Corticosterone, Sleep, Sleep deprivation, Sleep fragmentation, Rapid eye movement (REM), Rapid eye movement deprivation (REM deprivation)
Disrupted sleep is common and is widely considered to have adverse effects on general wellbeing and physical and mental health. Sleep disturbance interferes with diverse physiological processes such as metabolism [1] immune responses [2] neuroendocrine functioning, cognitive processing, and emotional states [3]. Moreover, loss of sleep often exacerbates preexisting conditions as diverse as hypertension [4] and bipolar disorder [5]. In animal models, REM deprivation can be devastating with disruption of long-term potentiation [6] induction of apoptosis [7] oxidative stress [8] and occasionally death [9].
Different types of sleep deprivation may have different physiological effects. Total deprivation of REM sleep (as might occur in a manic episode) is rare. Sleep is more commonly fragmented, separated into discrete blocks that may or may not include REM periods. Relative stress response between these has not been adequately examined. Using mice, we modeled REM deprivation using an adapted version of the inverted flowerpot method (IFM) [10] and modeled REM fragmentation using the moving bar method (MBM) [11] .
Male Black Swiss mice were obtained (The Jackson Laboratory, Bar Harbor, ME) and were housed in our animal care facility on a 12:12 light-dark cycle (6 am-6 pm). Mice were given one week to acclimate to the care facility. Protocols were approved by the Institutional Animal Care and Use Committee (Protocol Number 11078).
Sleep fragmentation chambers (Lafayette Instruments, Lafayette, IN) were utilized for REM fragmentation. Each cage contained 3-5 mice, with food and water available ad-libitum. Chambers were programmed so that a moving bar would sweep across the cage floor at 15 second intervals for one day (24 hrs) or three days (72 hrs).
For REM deprivation we utilized water-filled cages filled to 1 cm below the lip of a single platform (4 cm diameter). Food and water were provided for 30 min. three times daily. A human observed the animals continuously to rescue them if they fell into the water.
Control mice were housed in the animal care facility before being sacrificed. All mice were decapitated in AM for blood collection (between 8 AM and 2 PM, because it has been shown that corticosterone levels are stable over this time period in control mice [12]. After the serum was separated from the blood, it was stored in plastic tubes at -80 ℃. Corticosterone measurements utilized Arbor Assays Detect X® Enzyme Immunoassay kit (Catalog Number K014, Arbor Assays, Ann Arbor, Michigan) following manufacturer's protocol. Briefly, 1 µL of serum was treated with 1 µL of dissociation reagent for 5 minutes and diluted to 1:200. Samples were run in duplicate with a sheep polyclonal antibody against corticosterone. A microplate reader (Anthos Labtec Instruments, Salzburg, Austria) measured intensity at measurement filter 450 nm, and reference filter 405 nm for standards and all samples. Standard curves were created using My Assays.com online software (http://www.myassays.com/) and used to determine corticosterone concentrations of experimental samples.
Data were stratified according to intervention and duration of deprivation and compared with student t-tests. Proportions were examined by a test for proportional data [13]. A P value of < 0.05 was considered to indicate statistical significance.
Corticosterone levels were elevated compared to baseline in all conditions (Figure 1). Baseline level was 65.71 ± SEM 11.45 ng/mL in control mice (n = 18). REM fragmentation resulted in elevations of corticosterone concentrations after one day (148 ± 17.9 ng/mL, n = 11, t = 4.08, P = 0.0004), and three days (143.0 ± 22.4 ng/mL, n = 8, t = 3.42, P = 0.002). REM deprivation also increased corticosterone levels after one day (163.0 ± 13.8 ng/mL, n = 13, t = 5.42, P < 0.0001), and three days (147.6 ± 19.8 ng/mL, n = 12, t = 3.67, P = 0.001). Inter-group comparisons showed no difference between the different forms of sleep disruption (1-day t = 0.65, P = 0.52, 3-day t = 0.0143, P = 0.89).
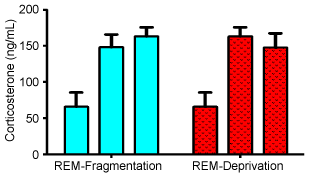 Figure 1: Serum corticosterone levels at baseline (1st column in each group), 1 day of treatment (2nd column in each group) and 3 days of treatment (3rd column in each group) for animals that underwent REM-fragmentation, and REM-deprivation. All treatment levels are significantly greater than baseline (p < 0.01), but none are different from each other. All values are means in ng/mL ± standard deviation. View Figure 1
Figure 1: Serum corticosterone levels at baseline (1st column in each group), 1 day of treatment (2nd column in each group) and 3 days of treatment (3rd column in each group) for animals that underwent REM-fragmentation, and REM-deprivation. All treatment levels are significantly greater than baseline (p < 0.01), but none are different from each other. All values are means in ng/mL ± standard deviation. View Figure 1
REM deprivation was more problematic than REM fragmentation. None of the animals died in the moving bar cages, but 3 mice expired in the 3-day inverted flowerpot group (0/8 vs. 3/22, z = 1.86, P > 0.5 [an additional 7 animals, 31.8%, did not die but had to removed from the experiment due to inability to remain on the platform before the 3 days were completed, z = 3.2, P < 0.01]).
Models of sleep disruption are usually associated with some other stressor that complicates interpretation [14-16]. Thus, elevations of corticosterone have been reported in many of the available animal models [17,18]. However, one of the major variables in these models is also the type of sleep disruption. Thus, the IFM is known to severely deprive animals from REM sleep [10] while the MBM allows animals to go into REM but fragments those periods [11]. We undertook this experiment because our experience was that REM-deprivation appeared to affect the animals more severely than REM fragmentation.
The results suggest that the IFM is more taxing on the mice than the MBM. Nearly 13.6% of animals exposed to the IFM died after 3 days, but none of the animals died in the MBM. This may be the first report of death in REM deprivation in mice [19] although REM deprivation appears to kill rats [9] and may play a role in unexplained deaths of human infants [20]. Furthermore, an additional 31.8% of mice exposed to the IFM could not complete the 3 days of the experiment, but all of the animals exposed to the MBM did complete the trial (P < 0.01). Thus, it is very surprising that there were no differences in corticosterone levels between the 2 methods at the end of the study (although the excluded animals were not examined) (Figure 1). Equally as unexpected is the absence of difference between 1 day of IFM (where all the animals completed the trial), and 3 days of the same procedure (where a total of 45.5% of the animals could not complete the experiment due to fatigue or death (Figure 1)). Our baseline measures of mouse corticosterone were in line with previous studies [11]. Thus, while we initially hypothesized that there would be a differential stress response between REM deprivation and REM fragmentation, the results of our study suggest no difference in corticosterone response.
There are important limitations to this present study to note. Historical [10] and recent studies [21,22] of sleep deprivation used rats. We used mice. Additionally, we did not monitor EEG to confirm the expected sleep disturbance of the two methods. The inverted flowerpot method is generally seen as more stressful to the mice due to social isolation, possible hypothermia, and relative immobilization [11,14]. In our studies we observed an excess of deaths and intolerance to the inverted flowerpot. This may have eliminated animals that would have had a greater corticosterone response, and brought the final average down. Finally, we did not examine other markers of stress response which may explain the differential response regarding death or intolerance between the two methods.
Despite these shortcomings, our present study presents evidence of similar HPA axis activation between REM fragmentation and REM deprivation. Yet other studies have noted that for short durations of sleep fragmentation there is no significant increase in cortisol after 6 hours [11]. There is a paucity of studies examining long duration sleep disturbance. Our study examined 24 and 72 duration of two different forms of disturbing sleep. While 3 days of exposure to IFM, but not MBM, was fatal in some animals, corticosterone response was equal in both groups.
Dr. Gao oversaw all aspects of this study including the performance of the work and the statistical analysis. Dr. Akers performed approximately half of the experimental work and the majority of the statistical analysis. Mr. Roberts performed the other half of the experimental work and contributed to the data analysis. Dr. El-Mallakh designed the experiment, oversaw most of the research and analysis, and wrote most of the paper.
Dr. El-Mallakh receives funding from the National Institute of Mental Health, the State of Kentucky, Merck, Teva, and Psychnostics. He is on the speakers bureau for Allergan, Merck, Lundbeck, Otsuka, and Takeda. None of the other authors have any conflicts of interest to report.
This work was performed without any extramural funding. Author El-Mallakh's other research is supported by the National Institute of Mental Health, the State of Kentucky, Merck, Teva, and Janssen. He serves on the speakers' bureaus of Allergan, Lundbeck, Merck, Otsuka, and Takeda. None of the other authors have any significant conflicts of interest to report.
Yonglin Gao supervised and performed the experiments, she supervised and performed the data analysis and also helped in the writing of the paper.
Brandon Akers performed the inverted flowerpot and some of the moving bar sleep deprivation experiments and performed much of the data analysis.
Michael B. Roberts performed the moving bar and some of the sleep deprivation experiments and helped Dr. Gao with the corticosterone measurements.
Rif S. El-Mallakh supervised all aspects of the experiments. He designed the experiments. He co-wrote the paper with Dr. Gao.