AMD3100 (Plerixafor), a specific antagonist of CXCR4, is the most potent small molecule non-peptide inhibitor to CXCR4/CXCL12 axis. The chemokine receptor CXCR4 and its ligand CXCL12 (SDF-1) expressed in a variety of tumor cells play an important role in regulating tumor biological behavior. The tumor microenvironment (TME) is the environment around a tumor, comprising blood vessels, immune cells, fibroblasts, signaling molecules and the extracellular matrix which are involved in tumor growth, invasion, metastasis, immune escape and tumor eradication. Although AMD3100 has been intensively investigated in tumor biology, it remains unclear how this treatment regimen modulates immune cells in the TME, which in turn affects the antitumor efficacy of other therapies. In this review, we specifically revisit the evidence from our and others' studies that AMD3100 acts as an immunomodulator to regulate immune responses in the TME and provide the perspective of synergy of AMD3100 with other therapeutics to prevent tumor development, progression, and metastasis.
CXCR4, AMD3100, Plerixafor, Tumor, The tumor microenvironment
Cancer is one of the leading causes of morbidity and mortality worldwide [1]. The high mortality rate of cancer is mainly related to tumor recurrence, metastasis and lack of effective treatment [2]. Increasing evidence suggests that the interaction between tumor and stromal cells is an integral part of the development and progression of tumors. The tumor microenvironment (TME) is best conceptualized as the composition of a variety of cells, such as noncancerous fibroblasts, adipocytes, immune and vascular cells, as well as signaling molecules and mediators [3-5].
It is gradually recognized that chemokines and their receptors are key communication bridges between tumor cells and stromal cells, creating a favorable microenvironment for tumor growth and metastasis. Chemokine 12 (chemokine 12, CXCL12), also known as stromal cell-derived factor-1 (SDF-1), is commonly expressed in many normal tissue cells and cancer cells [6]. In adult individuals, CXCL12 is involved in different physiological and pathological processes: Maintaining tissue balance; participating in the survival and recruitment of immune cells; and promoting the invasion and migration of certain tumor cells [7,8]. CXCR4 is a seven-pass transmembrane G protein-coupled receptor that is highly expressed in a variety of cells and certain tumor cells (such as breast, lung, and prostate cancer) and is associated with early metastasis and poor prognosis [9-11]. Tumor cells with high expression of CXCR4 bind CXCL12-expressing stromal cells and thus promote migration and invasion of tumors [7]. Activation of the CXCL12/CXCR4 axis promotes tumor cell proliferation, invasion, distant metastasis and inhibits various signaling pathways of apoptosis [12]. Therefore, the CXCL12/CXCR4 signaling pathway has become a very promising target for tumor therapy [13,14]. AMD3100, an antagonist of CXCR4, blocks the signaling of CXCR4 after binding to CXCR4, although cross-reactive to other chemokine receptors as previously reported [15], and thus play an important role in inhibition of tumor growth and metastasis. Emerging evidence demonstrates that the accumulation of immunosuppressive cells in the TME greatly reduce antitumor efficacy of the treatment, making it one of the major obstacles to cancer immunotherapy [16,17]. Immune cells are involved in different periods of tumor progression. In the initial stages of tumor development, macrophages directly kill tumor cells to promote antitumor responses and indirectly recruit and activate strong infiltration of other immune cells [18]. An intense infiltration of CD8+ T cells is usually associated with a good prognosis [19]. The presence of M2-polarized macrophages is widely recognized as a marker of poor prognosis [20,21]. Therefore, an important cancer treatment strategy is to regroup and redirect immune responses in the TME for a long-term elimination of tumor cells [2,22,23]. In this review, we focus on the regulation of immune cells by AMD3100 in the TME and detail four types of immune cells: CD8+ T cells, regulatory T cells (Tregs), tumor-associated macrophages (TAMs), and myeloid-derived suppressor cells (MDSCs).
The occurrence and development of tumors are inseparable from the TME. AMD3100 not only directly targets the CXCR4/CXCL12 axis to inhibit tumor growth and metastasis but also acts as a potent immunomodulator to enhance antitumor immune responses and prevent the development of a multi-faceted immunosuppressive intratumoral microenvironment (Figure 1).
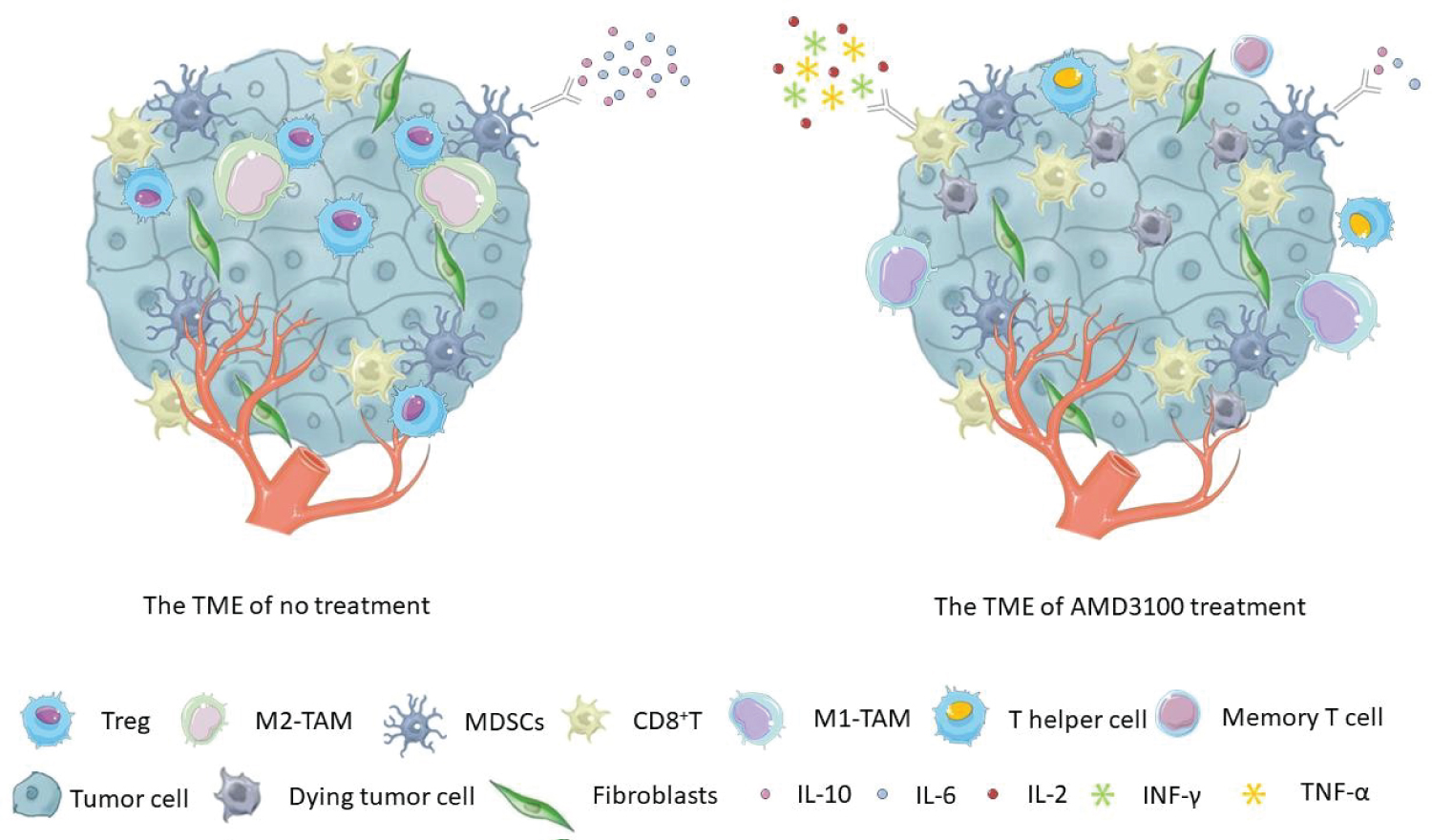 Figure 1: The tumor microenvironment (TME) was altered after treatment with AMD3100. It increased the proportion of CD8+ T cells and CD8+ memory T cells and promoted the conversion of Tregs into T helper-like cells in tumors. When used in combination with other therapies, AMD3100 enhanced the production of IFN-γ, TNF- α, or IL-2 in CD8+ T cells. The combination treatments also facilitated the polarization of M2 macrophages into M1, decreased the proportion of MDSCs or reduced the production of the immunosuppressive cytokines, such as IL-10 and IL-6.
View Figure 1
Figure 1: The tumor microenvironment (TME) was altered after treatment with AMD3100. It increased the proportion of CD8+ T cells and CD8+ memory T cells and promoted the conversion of Tregs into T helper-like cells in tumors. When used in combination with other therapies, AMD3100 enhanced the production of IFN-γ, TNF- α, or IL-2 in CD8+ T cells. The combination treatments also facilitated the polarization of M2 macrophages into M1, decreased the proportion of MDSCs or reduced the production of the immunosuppressive cytokines, such as IL-10 and IL-6.
View Figure 1
CD8+ T cells: In the TME, CD8+ T cells play an important role in antitumor immunity. CD8+ T cells differentiate into cytotoxic T cells (CTLs) and exhibit cytotoxicity against tumor cells. Terminally differentiated effector CD8+ T cells are IL-2 dependent and highly cytotoxic [24]. The overall survival rate of high-grade gliomas (HGG) was associated with the changes of tumor-infiltrating CD8+ T cells during treatment. AMD3100 in combination with bevacizumab resulted in a significant increase of CD8+ T cells [25]. In a pancreatic ductal adenocarcinoma (PDA) model, AMD3100 induced rapid accumulation of T cells in tumors and synergized with αPD-L1 to produce a significant reduction of tumors [26]. In addition, Zeng Y, et al. found that combination therapy with AMD3100 and αPD-1 further increased the infiltration of CD8+ T cells in ovarian tumors compared to αPD-1 monotherapy [27]. Some evidence suggests that AMD3100 increases the proportion of CD8+ memory T cells and enhances antitumor activity [27,28]. Furthermore, AMD3100 monotherapy and AMD3100-VIC-008 combination therapy inhibited PD-1 expression on CD8+ T cells in spleens, lymph nodes, and tumors of a mouse model of mesothelioma [29]. AMD3100, when combined with other therapies, significantly enhanced the expression of IFN-γ, TNF-α or IL-2 in CD8+ T cells [27,30].
Regulatory T cells: Regulatory T cells (Tregs) are immunosuppressive cells in lymphoid immune cells that interfere with antigen presentation by tumor cells and help tumor cells achieve immune escape. They are in a low number in normal tissue but significantly increase in many tumor tissues. Blocking the migration of Tregs into the TME is a promising approach for tumor immunotherapy [31]. Allogeneic hematopoietic cell transplantation (AlloHCT) has been supported as an effective treatment option for patients with refractory or relapsed non-Hodgkin's lymphoma. Dürr C, et al. elucidated that CXCR4 blockade not only reduced Treg migration into tumor tissue, but also enhanced the antitumor response after AlloHCT [32]. In addition, blocking CCL3-CCR1/CCR5 and CXCL12- CXCR4 axes inhibited Treg accumulation in the leukemic hematopoietic microenvironment (LHME) and delayed leukemia development [31]. Righi E, et al. first revealed that AMD3100 treatment impaired Treg migration in vitro and reduced CD4+ CD25+ and CD4+ CD25+ FoxP3+ T cells in vivo [33]. Li B, et al. further reported that Tregs were converted to IL2+ CD40L+ T helper-like cells and might lose their immunosuppressive function after the combined treatment with AMD3100 and VIC- 008 [29]. Following this study, Zeng Y, et al. further intensively studied the effect of AMD3100 on Treg cells and found that combination therapy with αPD-1 significantly inhibited Treg cells in tumors and increased the proportion of CD4+CD25- Foxp3+ T cells, phenotypically characteristic as CD40L+ IL-2+ T helper-like cells [27].
Tumor-associated macrophages: Tumor-associated macrophages (TAMs) are the most predominant and highly infiltrating cells in the TME [34]. TAMs regulate immune cell recruitment and inhibits antitumor responses by producing growth factors, chemokines and cytokines and resulting in an immunosuppressive TME [35]. TAMs are categorized as M1 or M2 types. M1 expresses proteins such as tumor necrosis factor (TNF), inducible nitric oxide lyase and MHC-II, which mainly enhance anti-tumor. M2 highly expresses CD163, CD204 and CD206, which promote tumor development [36]. AMD3100 has been demonstrated to inhibit M2 accumulation by blocking CXCR4/SDF-1 signaling [37]. The upregulation of CXCR4 in radioresistant tumor ECs is highly correlated with SDF-1TAM recruitment and TAM M2 polarization [38]. Recently, Zeng Yang, et al. showed that AMD3100 in combination with PD-1 reduced inhibitory leukocytes and promoted polarization to M1 from M2 macrophages in tumors in an ovarian cancer model [27].
Myeloid-derived suppressor cells: Myeloid-derived suppressor cells (MDSCs) are a group of immature bone marrow-derived cells with immunosuppressive functions. MDSC accumulation in tumors promotes immune escape [38,39]. Osteosarcoma tissues were infiltrated with a large number of MDSCs, which inhibit the expansion of CTLs. The majority of tumor-infiltrating MDSCs were CXCR4 positive and migrate toward the SDF-1 gradient. Binding of SDF-1 to its receptor CXCR4 activates the downstream AKT pathway, resulting in reduced apoptosis of MDSCs. And on this basis, AMD3100 promoted CD8+ T cell expansion by reducing CXCR4-expressing MDSCs in K7M2 osteosarcoma tumors [40]. The combined treatment with AMD3100 and αPD-1 significantly decreased the proportion of MDSCs and reduced the production of immunosuppressive cytokines IL-10 and IL-6, which might decrease the immunosuppressive effect [27]. Transmembrane TNF-α promotes the recruitment of MDSCs to tumor tissues by up-regulating the expression of CXCR4, but this can be partially blocked by treatment with the CXCR4 inhibitor AMD3100 [38]. The infiltration of MDSCs into the metastatic liver tumors may be mediated by up-regulating the expression of CXCR4, and AMD3100 blocks the interaction of tumor and stromal cells in the metastatic cascade to the liver [41]. Estrogen plays an important role in the occurrence, metastasis and drug resistance of estrogen receptor (ER) positive breast cancer. Ouyang L, et al. reported that after treatment with estrogen high levels of SDF-1α and tumor-infiltrating MDSCs were detected while blockade of the SDF-1/CXCR4 axis with AMD3100 neutralized the effect of estrogen on tumorigenesis [42].
The application of AMD3100 in cancer treatment has not been well characterized, particularly in AMD3100-mediated immune modulation. While effector T cells execute antitumor activities and eliminate tumor cells, immunosuppressive cells participate in tumor immune escape through various mechanisms. In-depth study of immune cells in the TME can provide useful diagnosis, prediction and prognostic information. The research of AMD3100 on the regulation of immune cells has profound significance and lays a solid foundation for future research and treatment, highlighting AMD3100 working synergistically with other therapeutics to enhance the antitumor efficacy.