Association between increased αβ-crystallin (ABC) and abnormal LN332 (LN332) expression in triple negative (TNBC) and basal-like (BLBC) breast cancers has been suggested recently. This study aims to evaluate ABC and LN332 immunohistochemical expression in a series of TN infiltrating ductal breast carcinomas and its association with clinical-pathological features and prognostic significance.
Microarrays of 69 cases of TNBC cases were scored for the expression of ABC, LN332, CK5/6 and EGFR by immunohistochemistry with linked clinical and pathological data (age, tumor size, lymph node metastases, histological grade and subtype, lymphovascular invasion). Tumors were scored as strong, moderate and weakly positive for these biomarkers.
ABC was expressed in 72.7% (48/66) and LN332 in 63.8% (44/69) of cases. ABC immunostaining was predominantly cytoplasmic (73.5%) and LN332 was exclusively cytoplasmic (100%). ABC was expressed in 83% of BLBC, followed by the penta negative profile (17%). The 5-year survival was 0.67 and 0.75 for ABC negative and LN332 positive tumors, respectively. For the BLBC group this probability was 0.67, while in the penta negative group (ABC, LN332, CK5/6 and EGFR) it was only 0.37. ABC-/LN332+ profile was associated with longer probability of non-recurrence (10-years probability: 0.86 for ABC-/LN332+ tumors versus 0.36 for other profiles; P = 0.021) and longer cumulative survival (10-years probability: 1.0 for ABC-/LN332+ tumors versus 0.53 for other profiles; P = 0.031). There was no association between the ABC-negative/LN332-positive profile and BLBC.
ABC and LN332 are commonly expressed in TNBC, especially in BLBC. TNBC with ABC-/LN332+ profile showed better clinical outcome.
Breast cancer, αβ crystallin, Laminin-332, Triple negative breast cancer, Basal-like breast cancer
Triple negative breast cancers (TNBC) show aggressive behavior and identification of new molecules for targeted therapies, as αβ-crystallin and laminins are needed. αβ-crystallin is one of the most characteristic "basal markers" of TNBC [1-3], and a gene profiling study identified that overexpression of αβ-crystallin is more highly correlated with this tumor type (45%) than with non-basal-like tumors (6%) [4].
αβ-crystallin is a small heat shock protein that promotes apoptosis resistance through binding pro-apoptotic Bax, Bcl-xs and p53 polypeptides and preventing their translocation to the mitochondria [5], acting as a molecular chaperone to bind and correct intracellular misfolded/unfolded proteins such as vascular endothelial growth factor [6-11]. In pathological conditions, like heat stress induction, αβ-crystallin can be induced in cell culture [12-14]. In vitro experiments revealed that αβ-crystallin promotes cell mobility and invasion [4]. Moreover, αβ crystallin can also act as an oncoprotein since, in nude mice, it has the intriguing property to transform immortalized human mammary epithelial cells in invasive mammary carcinomas that have the same aspect as basal-like breast tumors [7,15].
αβ crystallin expression in basal-like breast tumors is associated with poor patient survival independently of other prognostic markers [12,14-16]. These effects are likely mediated by direct interaction with actin and extracellular matrix proteins, including laminin. Recently, in a cohort of women with breast cancer brain metastasis, αβ-crystallin expression in primary breast carcinomas was associated with poor overall survival and poor survival after brain metastasis, across all subtypes and also within TNBC cases [17].
Laminins, a family of extracellular matrix glycoproteins, are the major non-collagenous constituent of basement membranes. They have been implicated in a wide variety of biological processes including cell adhesion, differentiation, migration, signaling, neurite outgrowth and metastasis [18]. Laminins are composed of 3 non identical chains: laminin alpha, beta and gamma (formerly A, B1, and B2, respectively). Each laminin chain is a multi domain protein encoded by a distinct gene [19,20]. Laminin-332 is a large multi domain α3β3γ2 isoform molecule involved in cell adhesion and matrix assembly, playing an important role in cell migration and survival by activating many signal mediators through binding to integrin α3β1 or α6β4. The role of laminin-332 in cell survival or anoikis resistance has been studied mostly in epithelial tumor cells, although some data exist on laminin-332-dependent survival of keratinocytes during wound healing [21]. Laminin-332 has been identified in immortalized human breast epithelial cells while α3, β3, and ϒ2 laminin-332 mRNA expression was found to be markedly down-regulated or not present in a panel of breast tumor cell lines when compared with normal breast epithelial cell lines [22]. Immunohistochemistry studies of laminin-332 expression in surgical specimens of mammary carcinoma revealed that 70% of TNBC are positive for laminin-332 [23] and its high expression levels have been described in metaplastic breast carcinomas and in the interface zone of breast cancer [23,24].
Therefore, this study aims to evaluate Αβ-crystallin and laminin-332 immunohistochemical expression in a group of TN infiltrating ductal breast carcinomas, as well as its relation with clinical-pathological features (age of the patient, tumor size, lymph node metastases, histological grade and subtype, lymphovascular invasion) and prognostic significance.
The histological files of all TNBC - ER, PR and HER2 negative were retrieved from a single institution over a period of 5 years (2001-2006). Triple-negative phenotype was defined as less than 1% of cells in the tumor staining for ER, PR and by 0 or 1+ staining of HER2. The sample set included 69 patients. Three cases were excluded from the αβ-crystallin stained tissue microarrays (TMAs) due to lack/little residual tumor in the cores (n = 66). This study was submitted to and approved by the Ethics Committee of the Research and Postgraduate Group of the Hospital de Clinicas de Porto Alegre, Brazil (GPPG 110263). Patient detail and clinical information were retrieved from medical records, including patient's age at diagnosis and tumor size, histological type and grade determined according to the Nottingham criteria. Histological types were diagnosed according to the World Health Organization (WHO) criteria. Follow-up (months to death/) ranged from 2-305 months, mean 71.4. Complete medical and treatment information was not available for the entire group of cases. Data on the time between tumor resection and fixation was not available.
The presence of αβ-crystallin and laminin-332 protein in human breast carcinomas was tested by immunohistochemistry performed manually on formalin-fixed tissue microarray (TMA) five micron thick sections mounted on charged glass slides. One section from each tumor array was stained with Hematoxilin & Eosin and was reviewed to confirm representative tumor tissue was included. Mouse monoclonal antibody αβ-crystallin class IgG1 (Novocastra, Clone: G2JF, cat# NCL-ABCrys-512) and purified mouse monoclonal anti-Kalinin B1 class IgG1 (BD Biosciences, Clone: 17/Kalinin B1, cat#610423) were applied on sections quenched with 3% H2O2. Steam heat or hot water bath (95 ℃) for 30 minutes was used for epitope retrieval in Citric Buffer pH6.2 or Cell Conditioning Solution (Ventana, cat#950-124) followed by incubation with avidin-biotin blocking (cat#sp2001, Vector Laboratories Inc). The anti-laminin-332 antibody, marketed as Kalinin B1 antibody (dilution 1:100) and αβ-crystallin (dilution 1:300) prepared in Ventana Diluent and 5% whole horse suppressor serum) was applied to the tissue sections overnight in a humid chamber at 4 ℃. The sections were then incubated with 1:500 dilution of biotinylated horse anti-mouse IgG (Vector Laboratories Inc cat# BA2000) for 30 min, followed by incubation with Streptavidin/peroxidase Conjugate (Roche Diagnostic, Ref# 11089 153 001). Reactions were developed by 3, 3'-diaminobenzidine tetrahydrochloride (DAB) and the slides were counterstained with Harris modified hematoxylin. Results of immunohistochemical staining were scored by pathologists (MG and DU) unaware of the tumor tissue samples. An inclusive list of antibodies used for this study, including clones, dilutions, antigen retrieval methods and manufactures is presented in Table 1.
Table 1: Antibodies used in this study. View Table 1
TMA were scored for nuclear and cytoplasmic staining for αβ-crystallin and laminin-332 stains in a semiquantitative way: weak, moderate and intense positivity. A cut-off of 1% was used to define positivity of these 2 markers. All cytoplasmic staining (≥ 1% of positive cells) for CK5/6 marker and all membranous staining for EGFR (≥ 1% of positive cells) was considered as positive result. The authors were blinded to the clinical information and staining results of other markers.
Findings were analyzed using the statistical software SPSS for Windows v.18.0. Chi-square was used to test for association between categorical variables (biomarker expression, tumor grade, histological subgroups) and exact p values are reported. Time-to-event analyses were done for survival data (time from breast tumor diagnosis to death or censoring) and disease-free survival (time from the date of breast cancer diagnosis to date of recurrence or censoring). The cumulative survival probability was estimated using the Kaplan-Meier method and the log-rank statistics was used to test for the relationship of survival with αβ-crystallin and laminin-332 expression. The data are presented as the means and standard deviations or absolute numbers and percentages.
A total of 69 cases were included in the study. The patient demographic data and pathological characteristics are shown in Table 2. Consistent with a TNBC population, patients presented advanced large, high grade tumors: 74% (42/57) were T2-T3 tumors, 68% (38/56) were grade 3 and lymph node involvement was seen in 54% (31/57) of the patients; 95% of the tumors were invasive ductal carcinomas of no specific tumor type and 45% of the cases showed lymphovascular invasion.
Table 2: Patient and pathological characteristics of 69 triple negative breast carcinoma cases. View Table 2
By immunohistochemistry, αβ-crystallin was positive in 73% (48/66) and LN332 in 64% (44/69) of cases. A basal-like profile was seen in the majority of cases (83%) compared to the penta negative tumor subgroup (17%). Representatives of αβ-crystallin and laminin-332 positive tumors are shown in Figure 1. αβ-crystallin immunostaining was predominantly cytoplasmic (74%) but definitive nuclear staining was observed in 15% of the cases. Rates of expression observed using a semi-quantitative 4-tier system of intensity classification (0, 1+/weak, 2+/moderate, 3+/strong) showed moderate/strong αβ-crystallin and laminin expression in 56% (37/66) and 37% (26/69) respectively (Table 3). Within the 3+ staining group, strong and diffuse cytoplasmic staining of αβ-crystallin was observed. Laminin-332 immunostaining was exclusively cytoplasmic in all positive cases. Table 4 presents the frequencies of αβ-crystallin and LN332 expression and the basal/penta negative profiles in relation to the different clinico-pathological characteristics of this series of triple negative breast tumors. The expression of laminin-332 in all positive cases was higher in basal-like (70%) than in penta negative cases (33%) (P = 0.022; Table 5). No difference in αβ-crystallin expression was observed between basal-like and penta negative groups. Because our sample consisted of TMAs, we could not detect differences in the geographical distribution of tumors.
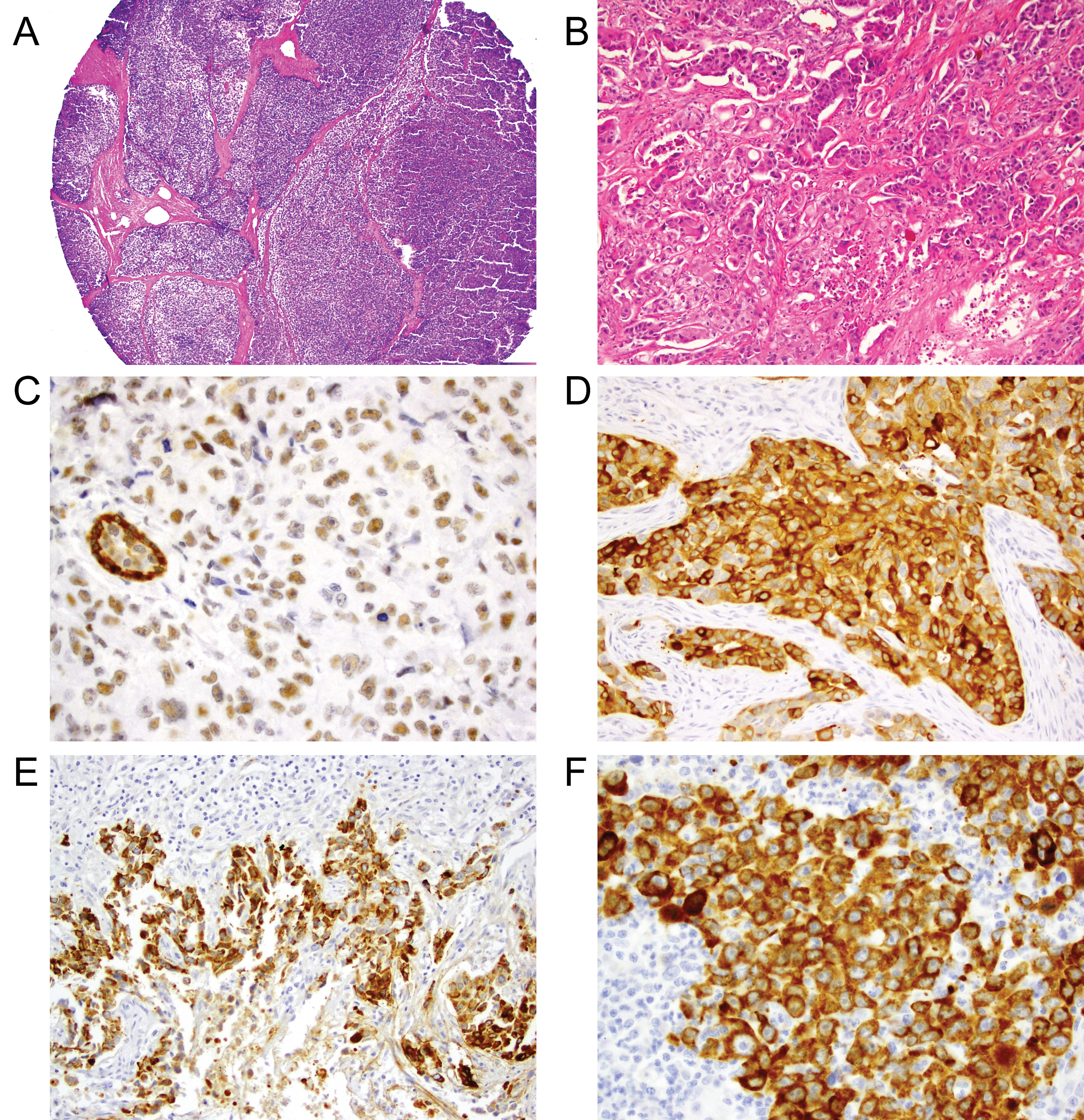 Figure 1: Examples of TMA core stained with H&E and the biomarkers.
Figure 1: Examples of TMA core stained with H&E and the biomarkers.
A) Example of TMA core on H&E stain (original magnification X 40); B) High grade TNBC on H&E (original magnification X 400); C) αβ-crystallin nuclear positive staining (original magnification X 400); D) Strong cytoplasmic αβ-crystallin expression (3+) (original magnification X 400); E) Laminin-332 strong cytoplasmic expression (3+) at tumor border (original magnification X 200); F) Laminin-332 cytoplasmic expression (original magnification X 200). View Figure 1
Table 3: Results of immunohistochemical semiquantitative scoring of αβ-crystallin and Laminin-332. View Table 3
Table 4: Frequencies of clinical-pathological characteristics according to immunohistochemical αβ-crystallin and laminin-332 positivity or basal/penta negative subtypes. View Table 4
Table 5: Relationships between immunohistochemical positivity for αβ-crystallin and laminin-332 and basal-like/penta negative profiles. View Table 5
Survival and tumor recurrence data were available for 55 patients. The cumulative probability of surviving > 60 months (5 years) for these women at the diagnostic date was 0.64 (Figure 2). When considering biomarkers expression, this probability was higher for LN332 positive (75%) compared to LN332 negative (42%) cases (p = 0.054) (see Table 4). On the other hand, the 5 year-cumulative survival was highest for cases with negative expression of αβ-crystallin (67% versus 55% for positive cases), but the difference was not statistically significant (p = 0.129). The basal-like group showed a 67% probability of surviving longer than 5 years (equal to the αβ-crystallin negative cases) and the survival for the penta negative group was 37% (p = 0.360). When the four combined profiles of αβ-crystallin/laminin-332 immunostainings (ABC+/LN332+; ABC-/LN332-; ABC+/LN332- and ABC-/LN332+) were compared, it was shown that the ABC negative/LN332 positive profile was associated with a statistically significant longer disease-free survival compared to the other profiles (Figure 3A; 10-year disease-free condition, 86% for ABC negative/LN332 positive tumors versus 36% for other profiles; p = 0.021). The ABC negative/LN332 positive profile was also associated with longer overall survival (Figure 3B; 10-year disease-specific survival, 100% versus 53% for other profiles; p = 0.031). There was no association between the ABC negative/LN332 positive profile and basal-like subtype.
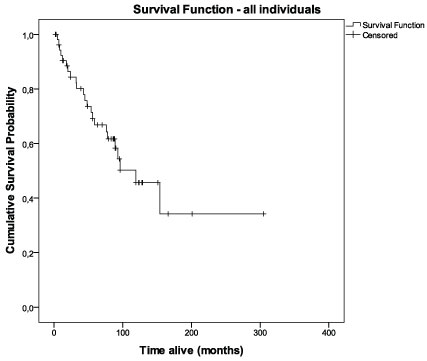 Figure 2: Cumulative survival curve for all cases (n = 69). View Figure 2
Figure 2: Cumulative survival curve for all cases (n = 69). View Figure 2
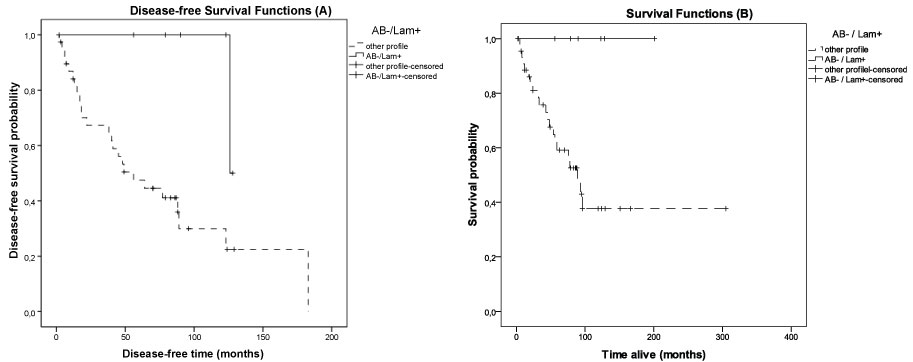 Figure 3: Cumulative probabilities of disease-free survival (A - left) and overall survival (B - right) for αβ-crystallin negative/laminin-332 positive profile compared to other profiles of expression for these biomarkers. View Figure 3
Figure 3: Cumulative probabilities of disease-free survival (A - left) and overall survival (B - right) for αβ-crystallin negative/laminin-332 positive profile compared to other profiles of expression for these biomarkers. View Figure 3
Our results confirm high expression of αβ-crystallin and laminin-332 in triple negative breast carcinomas, especially in the basal-like subtypes. The overall expression of αβ-crystallin in our series of triple negative breast cancer cases was observed in 73% of the cases, a higher value than that reported by Tsang, et al. in a series of 395 cases including 5 molecular subtypes of breast carcinoma (48%), but equal to the percentage of positivity reported in a series of breast cancer patients with brain metastases [12,17]. In a recent study, a significant positive association of αβ-crystallin expression was found with TNBC (48.6% sensitivity; 93.85% specificity) and basal-like carcinomas (56.5% sensitivity; 92.9% specificity) [25].
αβ-crystallin is one of the many heat shock proteins that function as inhibitors of apoptosis by preventing accumulation of denatured proteins and impedes the proteolytic cleavage activation of caspase-3. The overexpression of this molecule confers protection against a large panel of apoptotic stimuli and its silencing sensitizes cells to apoptosis. It has been associated with biologic aggressiveness and enhances cell survival in the setting of oncogenic stress, growth factor depletion, chemotherapy and other stressors [26-28]. Breast tumors that express this molecule have shown resistance to neoadjuvant chemotherapy and poor clinical outcome [29]. In an in vitro model of blood-brain barrier, αβ-crystallin promotes adhesion to brain microvascular endothelial cells, partly by a α3 β1 integrin-dependent mechanism. Previous work has shown that ABC binds to and corrects unfolded/misfolded VEGF under cellular stress and/or anti VEGF treatment [11]. It has been proposed as therapeutic target for TNBC treatment given its role in down regulating intracrine VEGF signaling. However, as direct therapeutic target, αβ-crystallin may cause deleterious side effects, given its protective role in cardiomyocytes and microvascular endothelial cells during ischemic heart disease [30]. A novel strategy to target αβ-crystallin using a small molecular inhibitor was shown to block breast cancer growth in vitro and an in vivo model [10].
We have also investigated the localization of αβ-crystallin staining characteristics and were able to demonstrate both cytoplasmic and nuclear IHC staining properties. The presence of αβ-crystallin in the nucleus has been described before in NIH3T3, NIE-115 and C2C12 cell lines under heat stress [13,31]. In normal cells αβ-crystallin seems to be predominantly found in the cytoplasm and translocates from the cytoplasm to the nucleus assuming an insoluble form upon stress. The αβ-crystallin speckles co-localize with intranuclear lamin A/C speckles upon heat, suggesting that this heat shock protein plays a role in formation and/or stabilization of the dynamic laminin-A/C architecture [13]. αβ-crystallin localizes in the nucleus with a speckled appearance, excluding nucleolar regions in C2C12 myoblast cell lines under heat stress [13]. In tumor tissue sections, αβ-crystallin nuclear staining was detected in samples of lung carcinomas (16%), cytoplasmic staining in 40%, and both nuclear and cytoplasmic staining positive in 10% of the cases [32]. There has been much interest in understanding the role of nuclear architecture in the regulation of nuclear functions [1].
Laminin-332 was identified in a high proportion (64%) of triple negative cases in this report. In most studies, laminin-332 has only been identified in immortalized human breast cell lines, but is either markedly decreased or not present in mammary carcinoma cell lines. Kwon, et al. tested the expression of LN332 in a series of surgically excised breast carcinomas by western blot and immunohistochemistry (IHC), and found a very similar 70% immunostaining in 80 triple negative breast carcinoma cases [23]. As described previously, laminin-332 is often more prominent in the cells at the tumor stroma interface, reminiscent of its basal location in normal breast [16]. The cytoplasmic immunostaining was specifically evaluated because it was more likely to be a product of the tumor itself rather than a secretory protein of stromal cells or residual basal epithelium.
Among the TNBC tumors, 57 out of 69 (83%) met a commonly used IHC definition of basal phenotype, namely EGFR and/or CK5/6 expression [33,34]. This percentage is comparable with the 76% of EGFR or CK5/6 staining tumors in a group of basal tumors defined by gene profile analysis. In this series of 57 basal-like tumors and 12 penta negative cases we could not show any differential αβ-crystallin expression, however the expression of laminin-332 was significantly higher in basal-like (70%) compared to penta negative cases (33%). αβ-crystallin has been described extensively in triple negative and metaplastic carcinomas, but laminin-332 expression has been less commonly reported. Laminin-332 has been identified in the basal layer of normal mammary gland, however a loss of the laminin-332 chains in more than 50% of the carcinoma stroma interface was reported in breast carcinoma cases [35]. Most of the studies show cytoplasmic staining in the invading border of carcinomas. Other reports showing downregulation of laminin-332 during tumorigenesis imply that it could be a potential marker for benign conditions. IHC studies show down regulation of laminin-332 in particular group of tumors like prostate carcinomas [36]. Additionally, downregulation of laminin-332 expression was identified in malignant and metastatic mammary carcinomas by gene transcription analysis. Laminin-332 mRNA was expressed at relatively high levels in MCF-10A immortal normal breast epithelial cells, long-term cultures of normal breast cells, and sorted primary cultures of normal breast luminal epithelial and myoepithelial cells. Reduced, but detectable, levels of laminin-332 tended to be expressed in cell lines derived from early-stage breast tumors, whereas expression was generally not detected in cell lines derived from later-stage tumors. In breast tumor tissue specimens, expression of laminin-332 α3 and β3 mRNAs tended to be reduced relative to levels observed in adjacent non tumor tissue, whereas gamma 2 levels were elevated in specimens with increased amounts of myoepithelial cells [22]. In contrast, our study provides evidence that laminin-332 and αβ-crystallin are often positive in one of the more aggressive subsets of mammary carcinoma. In a study of 243 cases of breast carcinoma including 80 triple negative carcinomas evaluated for laminin-332 IHC expression, 70% of TNBC carcinomas but only 15% of non-triple negative cases were positive [23]. Antibodies recognizing different epitopes of laminin-332 molecules were used in those studies and could explain the discrepant results. There is no study to resolve this discrepant laminin-332 production in particular groups of epithelial tumors.
The primary goal of this study was to describe αβ-crystallin and laminin-332 immunoexpression in a series of TNBC and to determine whether there is any association of their expression with prognosis. We were not able to show that the isolate expression of αβ-crystallin or laminin-332 affects overall survival rates. However, immunostaining of laminin-332, in the absence of αβ-crystallin positively affected prognosis and the reverse hold also true: immunostaining of αβ-crystallin in the absence of laminin-332 negatively affected prognosis. An earlier study showed a strong association with worse prognosis in Laminin-332 expressing hepatocellular carcinomas, in particular, the ϒ2 chain was strongly associated with the occurrence of metastasis [37]. The question remains whether laminin-332 expression has any modulatory effect on the migratory or invasive phenotype of breast carcinoma cells that show overexpression. A previous study on the co-expression of laminin-332 and its receptors, which might result in autocrine stimulation of migration, suggests this possibility [38]. Many reports describe the overexpression of heat shock proteins and extracellular matrix glycoproteins in cancer cells. This phenomenon is probably linked to the changes in protein homeostasis resulting of the accumulation of mutated proteins that promote cell migration and invasion by modulating signal transduction pathways in cancer [39]. Abrogation of tumor migration and progression induced by downregulation αβ-crystallin could be associated with repression of metalloproteinases concomitant with upregulation of tissue inhibitors, including matrix glycoproteins like laminin-332 [40]. This could be the explanation for the more "beneficial" αβ-crystallin-negative/laminin-332 positive profile suggested by our data.
In summary, our results underscore a previously unrecognized IHC signature for αβ-crystallin and laminin-332 in triple negative breast cancer cases and point to potential biomarkers to help identify patients within the triple-negative group that may present a less dismal prognosis. Further studies with genetic profiling of αβ-crystallin and laminin-332 in these tumors would help to clarify the biological relationship of these molecules in TN breast carcinomas.
This work was supported by a grants from Coordenação de Aprimoramento de Pessoal de Nivel Superior - CAPES (Bolsa Senior, processo 9437-13-4) and from Universidade Federal do Rio Grande do Sul (Edital 02/2012) and had financial support from Fundo de Incentivo a Pesquisa e Eventos (FIPE) do Hospital de Clinicas de Porto Alegre (GPPG 08446) and Department of Pathology of Memorial Sloan-Kettering Cancer Centre, New York.
This study was submitted to and approved by the Ethics Committee of our instituition and all the experiments comply with the current Brazilian laws.
The authors declare that they have no conflict of interest.