

International Journal of Pediatric Research
Single Center Cohort Study about in vitro Effects of Adalimumab and Anakinra on the Cytokine Network in Patients with Oligoarticular and Systemic Juvenile Idiopathic Arthritis
M. Kirchner*, B. Robinson, L. Strothmann, A. Sonnenschein and W. Mannhardt-Laakmann
University Hospital of Mainz, Department of Pediatrics, Division of Pediatric Immunology and Rheumatology, Mainz, Germany
*Corresponding author: Martina Kirchner, Department of Pediatrics, Division of Pediatric Immunology and Rheumatology, Langenbeckstrasse 1, D-55131 Mainz, Germany, Tel: 0049(0)6131-173963, Fax: 0049(0)6131-176491, E-mail: martina.kirchner@unimedizin-mainz.de
Int J Pediatr Res, IJPR-2-017, (Volume 2, Issue 1), Research Article; ISSN: 2469-5769
Received: February 03, 2016 | Accepted: March 25, 2016 | Published: March 28, 2016
Citation: Kirchner M, Robinson B, Strothmann L, Sonnenschein A, Mannhardt-Laakmann W (2016) Single Center Cohort Study about in vitro Effects of Adalimumab and Anakinra on the Cytokine Network in Patients with Oligoarticular and Systemic Juvenile Idiopathic Arthritis. Int J Pediatr Res 2:017. 10.23937/2469-5769/1510017
Copyright: © 2016 Kirchner M, et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Abstract
Background: Present data suggest that oligoarticular JIA is an autoimmune disorder with irregularity in both innate and adaptive immune responses, while sJIA shows distinct clinical and laboratory features, reflecting systemic inflammation. The activation of immunity stimulates the release of pro-inflammatory cytokines IL-1, IL-6 and TNF-α. Cytokine inhibition is considered as an appropriate therapeutic strategy for JIA. We investigated whether the blockade of a single cytokine pathway in the present cytokine setting causes an imbalance in the cytokine system.
Methods: This single center cohort study consisted of oJIA (n = 10) and sJIA (n = 10) affected children and control subjects (n = 25). We studied the cytokine secretion after LPS stimulation and in vitro inhibition of IL-1 and TNF-α by anakinra and adalimumab. Cytokine profiles from cell culture supernatants were examined with multiplex fluorescent bead immunoassay by flow cytometry.
Results: Adalimumab prevents highly effective and very selective the effect of TNF-α. Adalimumab and anakinra appear suppressive to IFN-γ. Anakinra unselectively inhibits the pro-inflammatory macrophage cytokines.
Conclusions: Our observations suggest that inhibition of IL-1 or TNF-α may contribute to the unselective decline of other pro-inflammatory cytokines in oJIA and sJIA patients. It still remains to be elucidated whether the reduced IFN-γ secretion is maybe causative for the increased susceptibility to infections with opportunistic pathogens.
Keywords
Oligoarticular Juvenile Idiopathic Arthritis, Systemic Juvenile Idiopathic Arthritis, Pro-/Anti-Inflammatory Cytokines, Cytokine Inhibition
Introduction
The oligoarticular juvenile idiopathic arthritis (oJIA) refers to a heterogeneous group of chronic inflammatory joint diseases of childhood and adolescence of unknown aetiology. The oligoarticular subtype is described as a T helper cell-mediated inflammatory disorder, mainly due the large number of activated Th1 cells and the abundance of pro-inflammatory cytokines in the inflamed joint [1]. This overproduction of pro-inflammatory cytokines is considered responsible for the clinical manifestations of JIA [2-4]. Th1 cells not only activate monocytes, macrophages and fibroblasts, but also B cells follow to produce immunoglobulins such as rheumatoid factor (RF) and antinuclear antibodies (ANA). In contrast, there are no signs of a T cell mediated, antigen-specific immune response in individuals with systemic-onset form of JIA (sJIA). The sJIA currently applies as auto-inflammatory disease, which goes back to an uncontrolled activation of the innate immune system [5]. The fact that modulation of pro-inflammatory cytokines results in improvement of clinical outcome of both JIA subtypes, underlines the important pathogenetic role of cytokines [6].
A primary role in the pathogenesis of oJIA play the macrophage cytokines interleukin-1 (IL-1) and tumor necrosis factor-alpha (TNF-α), which are detected in synovial fluids and or tissues of JIA patients, and are known to cause secretion of tissue-destructive matrix metalloproteinase. Furthermore, IL-1β and TNF-α inhibit production of matrix metalloproteinase inhibitors in synovial fibroblasts. This dual function favours joint damage [7]. Self-molecules, indicating synovial tissue damage, such as the calcium-binding proteins S100A8, S100A9 and S100A12 reinforce again the joint inflammation. Serum concentrations of S100A8/S100A9 proteins correlate very well with the disease activity in JIA patients, particularly sJIA [8,9]. Recent researches indicate that IL-1β is the prominent cytokine of sJIA. SJIA patients who did not respond to an anti-TNF-α treatment, showed an improvement in clinical and laboratory parameters after administration of IL-1 receptor blockers [10]. In addition to IL-1β and TNF-α, the cytokine IL-6 appears to be pathogenetically effective in the JIA. Circulating IL-6 is increased in all JIA subtypes and IL-6 serum levels of patients with increased disease activity correlate very well with their CRP levels [11]. IL-1β and TNF-α create a pro-inflammatory environment that favours the release of IL-17 from Th17 cells in the joint, leading to an additional increase in IL-6 levels. Particularly high IL-6 levels, which correlate well with the extent of joint involvement, can be found in the synovial fluid of sJIA patients [12]. The cytokine macrophage inhibitory factor (MIF) is a constitutive element of the host antimicrobial defenses and stress response that promotes pro-inflammatory function of the innate and acquired immune systems. MIF plays an important role in the pathogenesis of acute and chronic inflammatory or autoimmune disorders [13]. Herenius et al. have revealed that anti-TNF-α therapy diminishes serum levels of chemokine chemerin and thus inhibits the recruitment of MIF-producing cells (macrophages, dendritic cells), decreases MIF concentrations as well and eventually reduces inflammation in rheumatoid arthritis patients [14]. Endogenous MIF has been shown to exert a permissive regulatory role in cellular responsiveness to key pro-inflammatory cytokines TNF-α and IL-1 via up-regulation of cytokine receptor-dependent MAPK signaling independent of NF-κB [15].
The genetic basis of JIA is complex; the extent of associations with individual HLA genes varies greatly within the various subgroups. A few other genes, including macrophage inhibitory factor (MIF), TNF-α, IL-6 and IL-10 have also been discussed as disease relevant. Especially polymorphisms of the human MIF gene have been associated with increased susceptibility to or severity of juvenile idiopathic and adult rheumatoid arthritis. High amounts of IL-10 mRNA within the joint are associated with a milder and non-erosive onset form of oJIA [16,17].
Inhibition of IL-1β or TNF-α with soluble receptor fusion proteins or monoclonal antibodies have a proven effect on disease activity of the JIA. Anakinra is a genetically engineered human interleukin-1 receptor antagonist. It binds competitively to the IL-1 receptor without inducing a stimulatory signal [10]. Adalimumab is a human monoclonal antibody that binds to TNF-α and thereby an activation of the TNF-α receptor is prevented [18].
Notwithstanding the fact that the regulation of these cytokines results in a reduction of inflammation in patients with JIA, biologics are also responsible for a decreased immune response to infections which relates in particular those patients who are treated with IL-1 or TNF-α-blockers. Other possible side effects include the induction of autoimmunity and the occurrence of malignant tumors [19].
The persistent inflammation situation in JIA patients is most likely triggered by pathogen or damage associated molecular patterns (PAMPs, DAMPS). We therefore simulated this condition by LPS stimulation of leukocyte and investigated cytokines in oJIA-, sJIA patients and healthy after LPS stimulation and the addition of biologicals anakinra or adalimumab. In particular, we are interested in to which extent treatment with anakinra or adalimumab next to the desired anti-inflammatory effects may also affect the balance of the entire cytokine environment disadvantageous.
Methods
Patients and samples
This single center cohort study consisted of oJIA respectively sJIA affected children and control subjects and was conducted in the period 2009-2011. All JIA patients fit the ILAR classification criteria for childhood arthritis and provided written informed consent before enrolment. Peripheral venous blood was drawn from 20 patients of our pediatric rheumatological outpatient clinic and from 25 similar gender non-related young adults to serve as healthy controls. The study protocol was approved by the institutional ethics committee (#837.169.08).
Patient demographics
Ten oJIA patients and ten sJIA patients were enrolled after consent. Comprehensive clinical information was collected at each JIA patient visit, including history, physical examination (including presence of fever, rash and joint count), and clinical laboratory values [leukocytes, erythrocyte sedimentation rate (ESR), soluble IL-2 receptor (IL-2R), C-reactive protein (CRP) and complement activity (CH-50)]. Clinical status at each visit was graded according to a scoring system (Steinbrocker) to grade severity of arthritis. All patients were in stable remission or at least in controlled disease. Patients were under no therapy, antiphlogistic (Naproxene) and/or immunosuppressive (Methotrexate) therapy dependent upon their clinical symptoms. None of the patients was treated with immunosuppressive pharmaceutics used in our in-vitro model. Characteristics of the study subjects are shown in table 1.
![]()
Table 1: Patient demographics.
View Table 1
Isolation and culture of peripheral mononuclear cells (PBMCs)
Freshly isolated heparinized whole blood was overlaid on an equal volume of Lymphocyte Separation Medium (PAA-Laboratories GmbH, Pasching, Austria), and centrifuged at 800 × g for 30 min. The mononuclear cell ring, which contained PBMC, was obtained from the interface using a Pasteur pipette without disturbing the upper layer cells. The PBMCs were centrifuged again at 250 × g for 10 min. The pelleted cells were then resuspended in 5 ml phosphate buffered saline (DPBS, gibco, Paisley, UK). After washing, the pellets were suspended in RPMI 1640 medium containing 10% human AB-Serum, penicillin (100 U/ml), streptomycin (100 μg/ml) and glutamine (2 mM; all reagents from PAA, Germany). Next, 1 × 106 PBMCs/well were cultured in 200 μl of the RPMI 1640 for 24 h in round bottom 96 well plates (BRAND, Wertheim, Germany).
Co-culture of PBMC with gram-negative bacteria endotoxin, lipopolysaccharide (LPS)
PBMC samples (1 × 106/well) were incubated with 200 μl of RPMI 1640 with LPS (10 μg/ml; InvivoGen, San Diego, USA) at 37°C and 5% CO2. After 24 h of culture, the supernatants were obtained and kept at -20°C until use in the cytokine ELISA.
Inhibition of IL-1 or TNF-α in PBMC by anakinra and adalimumab
The BMC (1 × 106/well) were incubated with 200 μl RPMI 1640 containing LPS (10 μg/ml) and anakinra (1 μg/ml; Swedish Orphan Biovitrum AB, Stockholm, Sweden) or LPS (10 μg/ml) and adalimumab (10 μg/ml; Abbott Laboratories, Chicago, Illinois, USA) for 24 h at 37°C and 5% CO2. After 24 h, the supernatants were obtained and kept at -20°C until use in the cytokine ELISA.
Multiplex fluorescent bead immunoassay (ELISA) for determination of cytokine profiles
Two-colour flow cytometry was applied to cell culture supernatants of JIA patients and healthy control subjects, to investigate the concentrations of Interleukin (IL)-IL-1β, Interferon (IFN)-γ, IL-6, IL-10 and Tumor necrosis factor (TNF)-α. All cytokines were measured by commercial kit Human Th1/Th2 11plex FlowCytomix Kit (Bender MedSystems GmbH, Vienna, Austria) according to the manufacturer’s instructions for the use of tubes.
Statistical analysis
Mann-Whitney U test was used to compare data of healthy controls and JIA patients. Only the values of p < 0.05 were considered to be statistically significant in all analyses. Statistical analysis was performed with commercial software (SPSS Statistics Software version 23.0; SPSS Inc.).
Results
We examined the presence of twelve cytokines in leukocyte culture supernatants of 10 oJIA patients, 10 sJIA patients as well as 25 healthy individuals by flow cytometry analysis (multiplex fluorescent bead immunoassay). Selected results are shown in table 2.
![]()
Table 2: Median concentrations (pg/ml), minima and maxima of cytokines in patients with oJIA, sJIA and control subjects.
View Table 2
Pro-inflammatory macrophage cytokines IL-1, IL-6 and TNF-α
We stated that LPS stimulated leukocytes of oJIA patients secrete more IL-1 than leucocytes of sJIA patients or healthy individuals (Figure 1a). Upon LPS activation TNF-α secretion of oJIA leucocytes is much lower compared to control subjects and sJIA patients (Figure 1b). Interestingly, stimulated leukocytes of sJIA patients secrete more IL-6 than leukocytes of healthy individuals or oJIA patients (Figure 1c). Furthermore, in healthy subjects and sJIA patients a low inhibition of IL-1 could be achieved by anakinra, while oJIA patients showed changed IL-1 levels to a higher extent (Figure 2a). Adalimumab also reduces IL-1 levels, but only slightly and in oJIA patients and the control group (Figure 3a). By the use of adalimumab the complete inhibition of TNF-α was attained equally in both patient groups and healthy (Figure 3b). Although anakinra shows an inhibitory effect on TNF-α in in sJIA patients and healthy individuals it still seems to reinforce TNF-α secretion in oJIA patients (Figure 2b). No change in IL-6 concentration could be documented in oJIA patients during treatment with anakinra (Figure 2c). Healthy subjects and sJIA patients showed diminished IL-6 levels to a higher extent. While adalimumab has no significant effect on IL-6 in oJIA patients, it acts suppressive in sJIA patients and in the control group (Figure 3c).
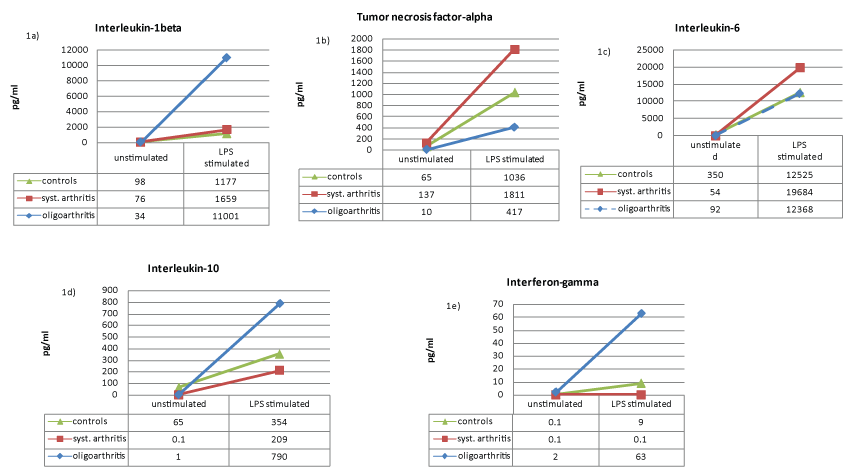
.
Figure 1: Connective dots showing the median cytokine values (pg/ml) before and after LPS stimulation of IL-1β (a), TNF-α (b), IL-6 (c), IL-10 (d) and IFN-γ (e) in healthy controls (green), sJIA patients (red) and oJIA patients (blue).
View Figure 1
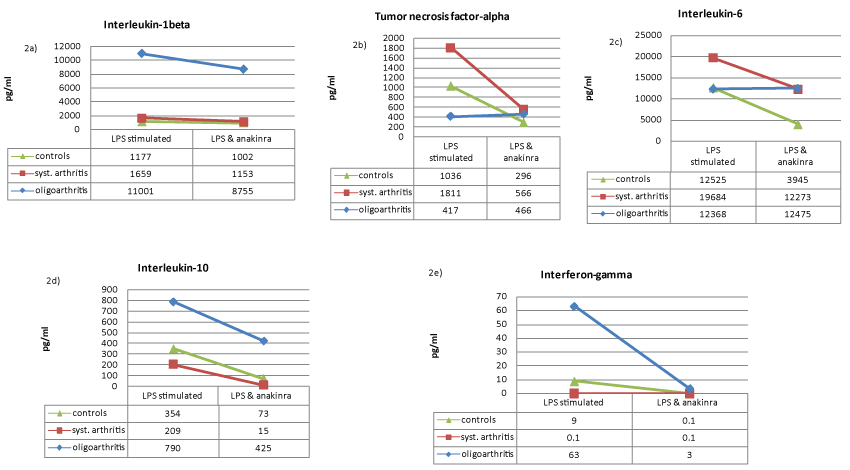
.
Figure 2: Connective dots showing the median cytokine values (pg/ml) under LPS stimulation before and after cytokine-blockade with anakinra of IL-1β (a), TNF-α (b), IL-6 (c), IL-10 (d) and IFN-γ (e) in healthy controls (green), sJIA patients (red) and oJIA patients (blue).
View Figure 2
Anti-inflammatory cytokine IL-10
LPS stimulated leukocytes of healthy individuals and oJIA patients secrete even more IL-10 than leucocytes of sJIA patients (Figure 1d). Furthermore, we found in sJIA patients as well as in healthy controls a severe reduction of IL-10 in stimulated leukocyte cultures with anakinra (Figure 2d). Anakinra seems to act less suppressive in oJIA patients. Adalimumab also reduces IL-10 levels significantly, but both patient groups showed diminished IL-10 levels to a lesser extent than the control group (Figure 3d).
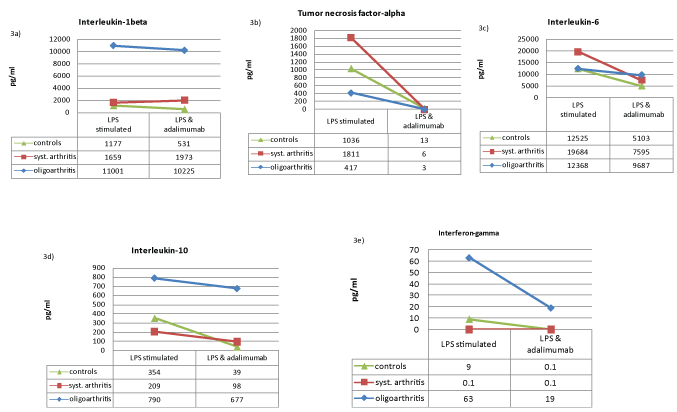
.
Figure 3: Connective dots showing the median cytokine values (pg/ml) under LPS stimulation before and after cytokine-blockade with adalimumab of IL-1β (a), TNF-α (b), IL-6 (c), IL-10 (d) and IFN-γ (e) in healthy controls (green), sJIA patients (red) and oJIA patients (blue).
View Figure 3
Pro-inflammatory T cell cytokine IFN-γ
Interestingly, stimulated leukocytes of healthy individuals secrete less IFN-γ than leukocytes of oJIA patients. We detected not any cytokine secretion of IFN-γ in sJIA patients (Figure 1e). We discovered that anakinra considerably inhibits IFN-γ in oJIA patients just as well as in healthy control subjects (Figure 3e). For adalimumab we observed a similar effect.
Discussion
A LPS-stimulation leads to increased expression of peptide/MHC complexes and T cell activation molecules on the surface of dendritic cells and activates this population over a MyD88- and TRIF-dependent signaling pathway. This signaling cascade induces clonal expansion and differentiation of CD4-positive T cells and thereby provides a link between innate and adaptive immunity. From our previous studies we know that JIA patients compared to healthy subjects show significantly higher concentrations of pro-inflammatory cytokines. In addition, we observed higher serum levels of spontaneous anti-inflammatory cytokine IL-10 in patients with oligoarticular JIA, seronegative arthritis and enthesitis associated arthritis [20].
During the present study, we were able to show that after LPS stimulation all groups studied show a pro-inflammatory cytokine profile, although to varying degrees. In oJIA patients outweighed the cytokines IL-1β and TNF-α. Our findings that LPS stimulated leukocytes of oJIA patients secrete more IL-1 than leucocytes of sJIA patients or healthy individuals (Figure 1a) differ from those of another research group. Pascual et al. showed in their study that PBMC of sJIA patients release large amounts of IL-1β upon activation with PMA-ionomycin [10]. One possible explanation for the varying results could be the fact that the types and amounts of cytokines produced may depend on stimulating reagents and treatments. It is known that different in vitro stimulations can have an effect on the results [21] and different stimulation protocols are often recommended by the manufacturer of reagents to obtain maximal cytokine production.
However, it is not ruled out that a sustained treatment with traditional synthetic disease modifying anti-rheumatic drugs affects cytokine levels. TNF-α production is stimulated by MIF which appears before TNF-α in the inflammatory cascade. Brennann-Bourdon et al. investigated in their cross-sectional study the association of MIF and TNF-α serum levels with methotrexate (MTX) and in combination with chloroquine (CLQ) and sulfasalazine (SSZ) in rheumatoid arthritis patients. They found that TNF-α serum levels were lower in the combination therapy group (MTX + CLQ + SSZ) in comparison with the monotherapy MTX group (16.7 pg/mL versus 13.6 pg/mL, p = 0.02). However, they did not find differences between DMARD therapies in MIF serum levels [22]. Therefore additional ex vivo data from patients using DMARD are required to clarify the effects of these drugs on cytokine levels.
However, in sJIA patients a significant increase in IL-6 has been shown. An enhanced IFN-γ production was found only at oJIA patients and healthy - not in sJIA patients. These results support the hypothesis that the cytokines TNF-α and IL-1β play a key role in the pathogenesis of oJIA and the cytokines IL-1β and IL-6 play a major role in the pathogenesis of sJIA. Lotito et al. also found higher concentrations of IL-1β at oJIA patients than in the control group [23]. In their study of the synovial fluid and synovial mononuclear cells of patients with oJIA and rheumatoid arthritis (RA) Caiello et al. analyzed the effects of IL-6 on the production of pro-inflammatory cytokines. They were able to show that IL-6 significantly increases the IL-1β secretion in combination with LPS or the endogenous Toll-like receptor ligands S100A8. The co-operation of IL-6 and DAMPS amplifies inflammation and leads to a positive feedback between the cells of the innate immunity and the stromal cells of the affected joints [24]. There, Th1-cells, activated monocytes and macrophages secrete IFN-γ, IL-1 is subsequently produced and therefore the synthesis of IL-10 inhibited [25,26]. Interestingly, after LPS stimulation we detected significantly less anti-inflammatory cytokine IL-10 in sJIA patients than in leukocyte cultures of oJIA patients and healthy individuals. The high effectiveness of in vitro stimulation of IL-1β could explain the intense suppression of IL-10 in sJIA patients.
Furthermore, in healthy subjects and patients with sJIA only a minimal inhibition of IL-1β could be achieved by anakinra, while in oJIA patients a stronger effect was observed (Figure 2a). Anakinra by itself -unlike anti-cytokine antibodies- acts as interleukin-1 receptor antagonist not directly on the concentration of the cytokine IL-1. Therefore unselective modulations of other cytokines are not unlikely. We observed a suppression of IL-6 (Figure 2c), TNF-α (Figure 2b) and IL-10 (Figure 2d) in healthy subjects and patients with sJIA. In contrast, in oJIA patients, a slightly elevated TNF-α secretion was documented. The reduction of IL-1β and TNF-α by anakinra apparently prevents the activation of CD4 positive T cells, and thus promoting the loss of IFN-γ secretion. The administration of anakinra or adalimumab resulted in a significant reduction of IFN-γ in the control group and in oJIA patients. Especially with anakinra a strong suppressive effect was observed regarding IFN-γ.
Adalimumab showed in the leucocyte cultures of healthy individuals and patients with oJIA also a suppressive effect on IL-1 secretion. For sJIA patients, this could not be shown (Figure 3a). The same applies to IL-6, however, the suppressive effect was not observed in oJIA- but in sJIA patients and healthy. This suggests that IL-6 after administration of anakinra or adalimumab plays a role similar to IL-1β in the sequence of inflammatory events. The complete inhibition of TNF-α by adalimumab could be demonstrated for all three study groups (Figure 3b). As a human monoclonal antibody adalimumab binds to TNF-α and directly prevents immediate and relatively selective efficacy of the cytokine. In oJIA patients high TNF-α levels normally lead as a result of increasing IL-12 and IL-18 production to an improved IFN-γ secretion [27]. After in vitro TNF-α-blockade the concentration of IFN-γ in patients and healthy individuals also fell off sharply. These results likely reflect the reduced macrophage activity. In addition, we observed a decrease in IL-10 secretion in the control group and in sJIA patients; in oJIA patients, however, the IL-10 concentrations remained nearly unaffected (Figure 3d). The therapeutic inhibition of inflammation via biologics is apparently not supported by up-regulation of anti-inflammatory cytokines.
Our studies on JIA patients show that the “selective” inhibition of a single cytokine influences the balance of the entire cytokine network. After administration of biologics the pro-inflammatory cytokines from macrophages - as intended - fell off in our experimental setup. However, as a side effect we observed also a significant decrease in the concentrations of the anti-inflammatory cytokine IL-10 and of the cytokine IFN-γ. This effect occurred in both anakinra as well as adalimumab. Several studies about clinical experience in use of biologics in children with JIA, have shown that biological agents can lead to clinical improvement only for a limited time, which is in accordance with our observation that an inhibition of a single cytokine can lead to a disturbance of the entire cytokine network, and therefore probably can lead to a disease flare. For example Lamot et al. assessed the long-term efficacy and tolerability of etanercept and infliximab in patients with JIA and observed that after initial improvement there is a flare in most of patients [28].
Conclusions
The results of our model confirm that TNF-α-antagonists are particularly suitable for the advanced treatment of oligoarthritis, while IL-1 antagonists are rather suitable for the treatment of systemic JIA. Although the use of biologics in pediatric rheumatology is considered relatively safe, our data help to understand how necessary is close monitoring with respect to unwanted and dangerous side effects. Studies from the adult rheumatology show that TNF-α -antagonists compared to other disease-modifying drugs pose an increased risk for opportunistic infections. Despite the intensive use of biologics in pediatrics there have been relatively few reports of serious infections in children and adolescents. According to BiKeR-index the infection rate is given as 2/100 patient-years for patients receiving etanercept [29]. It is unclear how the risk of infection for children displays at other biologics and what types of infections occur. However, so far there are only studies with a very low number of cases and the focus on treatment effectiveness for use of TNF-α-antagonists in polyarticular and systemic JIA. Bracaglia et al. reported that JIA-patients can frequently develop mild infections and, extremely rare serious infections when treated with adalimumab [30]. Beukelmann et al. showed in their studies that the JIA harbours an increased risk of infection independently from therapy [31,32]. Other long-term observations in the context of cohort studies or registries for the use of biologics will be able to provide evidence of the clinical utility of various treatment approaches and related security issues [29].
References
-
Berkun Y, Padeh S (2010) Environmental factors and the geoepidemiology of juvenile idiopathic arthritis. Autoimmun Rev 9: A319-324.
-
Kaminiarczyk-Pyzalka D, Adamczak K, Mikos H, Klimecka I, Moczko J, et al. (2014) Serum TNF-α levels and indicators of disease activity in children with oligoarticular juvenile idiopathic arthritis (oJIA) in the first year of the disease. Clin Lab 60: 799-807.
-
Muzaffer MA, Dayer JM, Feldman BM, Pruzanski W, Roux-Lombard P, et al. (2002) Differences in the profiles of circulating levels of soluble tumor necrosis factor receptors and interleukin 1 receptor antagonist reflect the heterogeneity of the subgroups of juvenile rheumatoid arthritis. J Rheumatol 29: 1071-1078.
-
Woo P (2002) Cytokines and juvenile idiopathic arthritis. Curr Rheumatol Rep 4: 452-457.
-
Martini A (2012) Systemic juvenile idiopathic arthritis. Autoimmun Rev 12: 56-59.
-
Beukelman T, Patkar NM, Saag KG, Tolleson-Rinehart S, Cron RQ, et al. (2011) 2011 American College of Rheumatology recommendations for the treatment of juvenile idiopathic arthritis: initiation and safety monitoring of therapeutic agents for the treatment of arthritis and systemic features. Arthritis Care Res 63: 465-482.
-
Mattey DL, Packham JC, Nixon NB, Coates L, Creamer P, et al. (2012) Association of cytokine and matrix metalloproteinase profiles with disease activity and function in ankylosing spondylitis. Arthritis Res Ther 14: R127.
-
Gerss J, Roth J, Holzinger D, Ruperto N, Wittkowski H, et al. (2012) Phagocyte-specific S100 proteins and high-sensitivity C reactive protein as biomarkers for a risk-adapted treatment to maintain remission in juvenile idiopathic arthritis: a comparative study. Ann Rheum Dis 71: 1991-1997.
-
Holzinger D, Frosch M, Kastrup A, Prince FH, Otten MH, et al. (2012) The Toll-like receptor 4 agonist MRP8/14 protein complex is a sensitive indicator for disease activity and predicts relapses in systemic-onset juvenile idiopathic arthritis. Ann Rheum Dis 71: 974-980.
-
Pascual V, Allantaz F, Arce E, Punaro M, Banchereau J (2005) Role of interleukin-1 (IL-1) in the pathogenesis of systemic onset juvenile idiopathic arthritis and clinical response to IL-1 blockade. J Exp Med 201: 1479-1486.
-
Havemose-Poulsen A, Sorensen LK, Stoltze K, Bendtzen K, Holmstrup P (2005) Cytokine profiles in peripheral blood and whole blood cell cultures associated with aggressive periodontitis, juvenile idiopathic arthritis, and rheumatoid arthritis. J periodontol 76: 2276-2285.
-
De Benedetti F, Pignatti P, Gerloni V, Massa M, Sartirana P, et al. (1997) Differences in synovial fluid cytokine levels between juvenile and adult rheumatoid arthritis. J Rheumatol 24: 1403-1409.
-
Renner P, Roger T, Calandra T (2005) Macrophage migration inhibitory factor: gene polymorphisms and susceptibility to inflammatory diseases. Clin Infect Dis 41 Suppl 7: S513-519.
-
Herenius MM, Oliveira AS, Wijbrandts CA, Gerlag DM, Tak PP, et al. (2013) Anti-TNF therapy reduces serum levels of chemerin in rheumatoid arthritis: a new mechanism by which anti-TNF might reduce inflammation. PLoS One 8: e57802.
-
Toh ML, Aeberli D, Lacey D, Yang Y, Santos LL, et al. (2006) Regulation of IL-1 and TNF receptor expression and function by endogenous macrophage migration inhibitory factor. J Immunol 177: 4818-4825.
-
Fossiez F, Banchereau J, Murray R, Van Kooten C, Garrone P, et al. (1998) Interleukin-17. Int Rev Immunol 16: 541-551.
-
Sugiyama E, Kuroda A, Taki H, Ikemoto M, Hori T, et al. (1995) Interleukin 10 cooperates with interleukin 4 to suppress inflammatory cytokine production by freshly prepared adherent rheumatoid synovial cells. J rheumatol 22: 2020-2026.
-
Imagawa T, Takei S, Umebayashi H, Yamaguchi K, Itoh Y, et al. (2012) Efficacy, pharmacokinetics, and safety of adalimumab in pediatric patients with juvenile idiopathic arthritis in Japan. Clin Rheumatol 31: 1713-1721.
-
Horneff G, Foeldvari I, Minden K, Moebius D, Hospach T (2011) Report on malignancies in the German juvenile idiopathic arthritis registry. Rheumatology (Oxford) 50: 230-236.
-
Kirchner M, Sonnenschein A, Schoofs S, Schmidtke P, Umlauf VN, et al. (2013) Surface expression and genotypes of Toll-like receptors 2 and 4 in patients with juvenile idiopathic arthritis and systemic lupus erythematosus. Pediatr Rheumatol online J 11: 9.
-
Baran J, Kowalczyk D, Ozóg M, Zembala M (2001) Three-color flow cytometry detection of intracellular cytokines in peripheral blood mononuclear cells: comparative analysis of phorbol myristate acetate-ionomycin and phytohemagglutinin stimulation. Clin Diagn Lab Immunol 8: 303-313.
-
Brennan-Bourdon LM, De la Cruz-Mosso U, Reyes-Castillo Z, Martínez-Bonilla GE, Ramírez-Dueñas MG, et al. (2015). MIF and TNFalpha serum levels in rheumatoid arthritis patients treated with disease-modifying antirheumatic drugs: a cross-sectional study. Immunopharmacol Immunotoxicol 37: 207-213.
-
Lotito AP, Campa A, Silva CA, Kiss MH, Mello SB (2007) Interleukin 18 as a marker of disease activity and severity in patients with juvenile idiopathic arthritis. J Rheumatol 34: 823-830.
-
Caiello I, Minnone G, Holzinger D, Vogl T, Prencipe G, et al. (2014) IL-6 amplifies TLR mediated cytokine and chemokine production: implications for the pathogenesis of rheumatic inflammatory diseases. PLoS One 9: e107886.
-
Gattorno M, Prigione I, Morandi F, Gregorio A, Chiesa S, et al. (2005) Phenotypic and functional characterisation of CCR7+ and CCR7- CD4+ memory T cells homing to the joints in juvenile idiopathic arthritis. Arthritis Res Ther 7: R256-267.
-
Agarwal S, Misra R, Aggarwal A (2008) Interleukin 17 levels are increased in juvenile idiopathic arthritis synovial fluid and induce synovial fibroblasts to produce proinflammatory cytokines and matrix metalloproteinases. J Rheumatol 35: 515-519.
-
Wedderburn LR, Robinson N, Patel A, Varsani H, Woo P (2000) Selective recruitment of polarized T cells expressing CCR5 and CXCR3 to the inflamed joints of children with juvenile idiopathic arthritis. Arthritis Rheum 43: 765-774.
-
Lamot L, Bukovac LT, Vidovic M, Frleta M, Harjacek M (2011) The 'head-to-head' comparison of etanercept and infliximab in treating children with juvenile idiopathic arthritis. Clin Exp Rheumatol 29: 131-139.
-
Horneff G, Minden K, Foeldvari I, Onken N, Windschall D, et al. (2014) [Current news from the BIKER register]. Z Rheumatol 73: 897-906.
-
Bracaglia C, Buonuomo PS, Tozzi AE, Pardeo M, Nicolai R, et al. (2012) Safety and efficacy of etanercept in a cohort of patients with juvenile idiopathic arthritis under 4 years of age. J Rheumatol 39: 1287-1290.
-
Beukelman T, Xie F, Chen L, Baddley JW, Delzell E, et al. (2012) Rates of hospitalized bacterial infection associated with juvenile idiopathic arthritis and its treatment. Arthritis Rheum 64: 2773-2780.
-
Beukelman T, Xie F, Baddley JW, Chen L, Delzell E, et al. (2013) Brief report: incidence of selected opportunistic infections among children with juvenile idiopathic arthritis. Arthritis Rheum 65: 1384-1389.
