

Journal of Toxicology and Risk Assessment
Micronuclei Induction and Neurotoxic Effect in C6 Glioma Cells Exposed to Low Concentrations of Diazinon, an Organophosphorus Compound
Carla Falugi1*, Chiara Gambardella2, Tommaso Bonfiglio3, Zoltan Rakonczay4, Federico Biggi1, Sara Novelli1 and Mariangela Masini1
1Dipartimento DISTAV, University of Genoa, Italy
2ISMAR, Consiglio Nazionale delle Ricerche, Italy
3DIFAR, University of Genoa, Pharmacology and Toxicology, Italy
4Department of Oral Biology and Experimental Dental Research, Faculty of Dentistry , University of Szeged, Szeged, Hungary
*Corresponding author:
Carla Falugi, Dipartimento DISTAV, Università di Genova, Viale Benedetto XV, N°5, CAP 16132 Genova (Italia), Italy, E-mail: carlafalugi@gmail.com
J Toxicol Risk Assess,
JTRA-1-001, (Volume 1, Issue 1),
Original Article
Received: July 23, 2015: Accepted: August 24, 2015: Published: August 26, 2015
Citation: Falugi C, Gambardella C, Bonfiglio T, Rakonczay Z, Biggi F, et al. (2015) Micronuclei Induction and Neurotoxic Effect in C6 Glioma Cells Exposed to Low Concentrations of Diazinon, an Organophosphorus Compound. J Toxicol Risk Assess 1:002
Copyright: © 2015 Falugi C. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Abstract
Background:The presence of doses of diazinon far lower than IC50 cholinesterase activity was reported in plasma of pregnant women and newborns living in agricultural areas.
Objective:In the current study, we investigated the possibility of neurotoxicity induction by exposing cultured gliotypic C6 cells to a similar range of concentrations, for 24 h, at 37°C.
Materials and methods:Confluent C6 cellswere exposed to diazinon (DZN) at concentrations from 200ng/L to 0.002ng/L. The maintenance of confluence, the induction of micronuclei and the expression of molecules related to the cholinergic system were verified, by morphological, biochemical and immunohistochemical methods, in order to check the effects of the altered modulation of the cholinergic signal on glial-like cells.
Results:The exposure to 0.002ng/L showed significant effecton micronuclei occurrence since the exposure to 0.002ng/L, while the inhibition on butyrylcholinesterase activity showed significant variations starting from the exposure to 0.2ng/L up to 200ng/L. Acetylcholinesterase activity was significantly inhibited only by the exposure to 200ng/L. The immunohistochemical localization of choline acetyltransferase and fibronectin showed dramatic variation only in C6 cells exposed to 200ng/L.
Conclusion:The low doses of DZN investigated affect the investigated features of glial-like cells, mainly starting from the 0.2ng/L dose, while the effects on AChE activity and ChAT and fibronectin-immuorectivity were clearly exerted in cell cultures exposed to DZN 200ng/L. Collectively, these findings translated to the in vivo functions of glial cells indicate that exposure to doses that are nontoxic to adult organisms may weaken the brain defense and functions of glial cells through an AChE-mediated mechanisms.
Keywords
C6 glioma cells, Choline-acetyltransferase, Cholinesterase activities, DZN, Fibronectin, Inhibition, Micronuclei, Neurotoxicity, Organophosphate insecticide
Abbreviations
ACh: Acetylcholine, AChE: Acetylcholinesterase (EC 3.1.1.7), BChE: Pseudocholinesterase or butyrylcholinesterase (EC 3.1.1.8), BSA: Bovine Serum Albumin, ChAT: Choline Acetyltransferase (EC 2.3.1.6), DAPI: 4'-6-Diamidino-2-Phenylindole, FN: Fibronectin, IR: Immunoreactivity, MN: Micronuclei, PBS: Phosphate Buffered Saline, PFA: Paraformaldehyde, PI: Propidium Iodide
Introduction
Even though the adverse effects on environment and organism health have been long and thoroughly studied, the problem represented by the continuous use of neurotoxic pesticides, in particular organophosphates (OPs, organic salts of the pyro phosphoric acid), is still open. These are considered as emerging pollutants [1], as their massive employment is relatively recent, dating after the ban of persistent organic pesticides. OPs are not persistent for a long time, as their half-life varies according to the climate and ground substrates, and generally is estimated of some weeks. Nevertheless, their continuous use along the year in large agricultural sites makes their presence stable in the environment. The mechanism of action is due to their ability to inhibit the activity of acetylcholinesterase and other key enzymes related to the modulation of neurotransmission [2]. The acute-dose exposure, generally due to accidentsduring agricultural operations, is able to cause a number of neurologic symptoms, up to death [3]. Moreover, a significant association between prenatal exposure to OPs and anomalous primitive reflexes was observed in newborns [4]. These involve a cluster of automatic and stereotyped movements, such as breathing, blinking, swallowing and glandular function, and are relevant for further development of the nervous system.
Among OPs Diazinon (DZN) was one of the most widely used insecticides for household and agricultural pest control. In 2000, the United States Environmental Protection Agency (U.S. EPA) announced an agreement with the registrants of DZN to cancel all residential uses of the pesticide, leaving only agricultural uses [5]. As currently the use is restricted to agricultural practice, the main way in which the public could be exposed is through eating food directly or indirectly treated with DZN, or by use of water where it is present as a result of run-off and movement through the soil from areas where it is used in farming [6].
Although very few reports are available in the literature about the amount of organophosphate inside human organisms, the presence of DZN has been found and measured in the plasma of women and newborns living in agricultural sites [7]. In addition, the presence of OP metabolites in the urine of children of preschooler age showed a day-by-day variation according to the daily dietary exposure, and an increase during the periods of orchard treatments [8,9]. Newborns and immature animals actually recover rapidly from cholinesterase inhibition [10] so measurements of cholinesterase activity other than in the immediate exposure period can give a misleading assessment of adverse effects. [10,11].
Inside the organisms of women and newborns living in agricultural sites, or feeding on non-organic food, DZN levels in plasma were reported to range between 0 and 0.5mg/L [5]. Which is much lower than the acute cencentrations. These doses are non-toxic for adult organisms, but inside embryos and newborns they are into direct contact with developing target cells. In this work, we used concentrations ranging between 0.002 and 200ng/L, in order to verify if stress and possible risk for neural health may be induced by DZNconcentrations falling within the described range present in plasma. To this aim, we used C6 rat glioma cultured cell line as a model of cells responsible for the brain defense. Actually, glial cells are part of the brain-blood barrier, formed by the association of brain microvessels, astrocytes and neurons, which form functional 'neurovascular units' [12]. These cells enter into contact with the substances present in the plasma. C6 cells derive from astrocytes, and retain most of their properties, e.g. the presence of a set of cholinergic molecules [13,14] besides a wide array of receptors, transporters and ion channels, allowing them to sense and dynamically modulate neuronal activity [15].
The C6 cells were exposed to to DZN concentrations mimicking the doses present in the blood of women and newborns, and the exposure lasted for 24 h, in order to check the scenario of a single-day exposure to the day-by-day variable amount, as above described [9,10]. In this short period bioaccumulationis not relevant, and oxonization needs a more complex system than non-chronically exposed cell cultures. Thus the results have been directly correlated to the changes of AChE activity, as determined by biochemical measurements.
Materials and Methods
The C6 cells (CCL-107) were obtained from American Type Culture Collection, Rockville, MD. The chemicals were obtained from Sigma Chem. Co, IT, unless otherwise stated. The purity of chemicals was verified according to the data sheet that accompanied the delivered materials; the decay time for each reagent was written on the bottles; the maintenance of the properties of the C6 cells was verified at each step by the biochemical and immunohistochemical controls. Culture media were from Gibco-Invitrogen, Karlsruhe, DE. Plasticware was from Falcon/Fred Baker, Runcorn, UK. Purified enzymes used for comparison and IC 50 assessment were Electric eel, rat brain AChE and purified mammalian plasma BChE, commercially obtained from Sigma. Purified Diazinon (DZN) was obtained from PESTANAL®, through Sigma (IT) purchase. The purity of DZN, according to the data sheet was 98%. DZN molecular weight is 304.4 [10,4]. The DZN Chemical Abstracts Service (CAS) registry number is 333-41-5. The CAS and IUPAC name is O, O-diethyl O-[6- methyl-2-(1-methylethyl)-4-pyrimidinyl] phosphorothioate [5,6]. A mother solution was performed by dissolving 2mg of DZN in 1 L H2O, without using other solvent, as DZN solubility in water is 40mg/L at 20°C. From this stock, solutions in the culture medium (200,0.2 and 0.002ng/L) were prepared and used to expose the C6 cell cultures for 24 h.
Exposure experiments
The exposure experiments were repeated three times with different pools of C6 cultured cells.Rat brain C6 Glioma cells were routinely cultured on sterile slides in RPMI 1640/5% NCS (heat-inactivated) and 2mM/L-glutamine in a humidified atmosphere of 5% CO2, up to confluence, and then exposed for 24 h at T=37°C to DZN at the different concentrations. Control samples were incubated at the same time without DZN addition.
Cultures morphology (non-fixed cells)
The general appearance of the unstained and stained cultures was observed on a light microscope (Nikon, Eclipse TS100).
Viability assessment: The viability of the cultured cells was examined by use of double staining. The cells were treated with DAPI (4',6-diamidino-2-phenylindole, GIBCO, IT) 1mg/L in 0.1M citrate buffer pH 7 for 10 min in the dark. Successively the cells were treated with 1mg/L RNAse (Sigma, Italy) for 30 minutes at 37°C and stained with 50mg/L Propidium Iodide (PI, BioChemika, Italy) for 5 minutes at room temperature. DAPI (4',6-diamidino-2-phenylindole) is a blue-fluorescent DNA stain that exhibits ~20-fold enhancement of fluorescence upon binding to dsDNA. It is excited by the violet (405nm) laser line and is commonly used as a nuclear staining of both living and dead cells [16]. PI selectively stains with red fluorescence the nucleic acids of dead cells, because it cannot pass through the living membranes. To prevent RNA staining, PI incubation was preceded by pre-incubation with RNAse. The cells stained with PI were counted and compared with the number of cells stained with DAPI, in order to establish the ratio of dead vs total number of cells.The cells were thenrinsed in clean incubation mediumand observedon a Leitz microscope equipped for epifluorescence appropriate filters.
Occurrence of micronuclei[17]: DAPI staining was also used to quantify the presence of micronuclei. We performed the micronucleus test as previously described [18], i.e. during the recovery after contaminant-induced cell death, shown by necrotic holes in the tissues.
For each exposure and controls we observed 6 slides (in total 24 slides). On each slide, we pointed the microscope objective40x on 6 casual fields (field=the part of the slide that is seen in the objective). For each field, containing different number of cells according to the exposures, we counted the number of micronuclei vs the total number of cells. Thus, for each exposure, we had 6 ratios. Of these, we calculated the average values and the standard deviations, and the significance according to One-Way Anova and Levene's from medians.
The observation was made by the Leitz microscope equipped with UV apparatus and emission filter for 470nm wavelength (Leitz, Germany).
Cholinesterase activities: The activity of both acetylcholinesterase (AChE, EC,3.1.1.7) and butyrylcholinesterase (BChE, EC 3.1.1.8) was measured by spectrophotometric method [19], which was modified ad hoc for the Jenway (6405 UV/vis) spectrophotometer.
The control and exposed cultured cells at 24 h exposure were collected and maintained for one night at -20°C. The frozen cells were then brought to room temperature, homogenated with a minipotter, passed through a syringe with thin needle, (Ultrafin 29G, 12,7mm length) in the presence of 1% triton X100, sonicated for 25min in a bath sonicator (FALC, mod. LBS1, Italy) and centrifuged for 3min at 8000rpm. The resulting supernatant was used for the spectrophotometric analyses. The kinetics of AChE and BChE activities were measured by the absorbance at λ=412 nm. The rate of substrate cleavage was compared with the standard curve that had been previously obtained by supplying known amounts of ChEs. Protein concentration was measured with the Coomassie Brilliant Blue protein-binding method [16], using bovine serum albumin as standard. (BSA, Sigma). The AChE units here reported corresponded to micromoles of substrates hydrolyzed/min/mg protein at room temperature.
The measurements were performed with cells from different cultures, in triplicate, and were repeated four times along the duration of the work.
Specificity controls of the ChE reaction were performed by using specific inhibitors: 10mM BW284c51 (1,5-bis(4-allyldimethylammoniumphenyl)pentan-3-one dibromide) and Iso-OMPA (tetraisopropyl pyrophosphoramide, both obtained from Sigma, Italy) in the reaction medium. BW284c51 is a specific inhibitor of the so-called "true AChE" (E.C.3.1.1.7), and Iso-OMPA is a selective inhibitor of the pseudocholinesterase (BChE, E.C. 3.1.1.8).
IC50 was established by the same methods, by use of purified enzymes (Electric eel and rat brain AChE, mammalian plasma BChE) obtained by Sigma. AChE activity was assayed in the presence of 0.1mM Iso-OMPA to inhibit BChE. The IC50 values of the inhibitors were calculated by linear regression of the log concentration versus inhibition functions (range 20-80% inhibition). Briefly, AChE was preincubated with DZN for 30 min and the reaction was started by addition of the substrate acetyl-β-methyl thiocholine iodide. All assays were performed in triplicate in 3-5 independent experiments.
At these high concentrations, DZN is not fully water soluble; MeOH (methyl alcohol) was used as a solvent. We previously tested the effect of MeOH on purified ChE activities, and found that the optimal final concentration in the incubation mixture of MeOH was 0.5%. We also found that minor concentrations of MeOH in some cases increased the inhibition efficiency of insecticides (data not shown).
Statistical elaboration of data
Statistical elaboration of data related to the enzyme activity and the related graphics were performed by comparable tests (One-Way Anova and Kruskal Wallis tests, by the PAST and Primer programs).
Immunocytochemical reactions
Fixation: The slides carrying the control and exposed C6 cell cultureswere fixed with 2% paraformaldehyde in phosphate buffered saline (PBS: phosphate buffer 0.1M, pH 7.4 with 0.8% NaCl) for 10 min at room temperature and prepared for microscopic analyses. The slides were rinsed in 0.1M PBS (pH 7.4), permeabilized by cold methanol and 20% polyethylene glycol (Sigma, Italy) and subsequently rehydrated for cytochemical and immunocytochemical reactions.
Cytological localization of molecules immunologically related (IR) to fibronectin (FN) and choline-acetyltransferase (ChAT, EC 2.3.1.6): Control and exposed slides were incubated overnight in a moist chamber at 4°C with the primary rabbit polyclonal antiserums to fibronectin (FN) and to choline-acetyltransferase (ChAT) (Dako, Denmark), respectively. The antisera were diluted 1:200 in PBS (pH 7.4) with 0.1% bovine serum albumin (BSA). The negative control was performed by omitting the primary antiserum. Aspecific reactions were also blocked by preincubation in cold PBS containing 0.5M glycine and 1% BSA and 5%normal goat serum (GSA). After incubation, samples were rinsed in PBS and incubated for two hours at room temperature in the secondary anti-rabbit Alexa 488 (Molecular probes, Invitrogen Corporation, USA) 1:200 in PBS. All samples were thoroughly washed in PBS, counterstained with PI as previously described and mounted on slides with the antifading agent Gelvatol [20,21] for microscopic observations.
Results
C6 cell responses were differently affected by the different concentrations of DZN; actually MN occurrence showed significant variation from controls already for exposures to the minor concentration of the pesticide (0.002ng/L); the morphology of the cultures showed an evident effect since the concentration 0.2ng/L, ChE and immunocytochemical staining showed evident effects after exposure to f the higher concentration of DZN (200ng/L).
Cultures morphology: Unfixed, unstained slidesFigure 1A shows the aspect of a confluent control sample after 24h. The cells appear very close each-other, transparent and without clusters. Figure 1B shows a slide carrying C6 cultured cells after 24h exposure to 200ng/L. The culture showed loss of confluence, huge portions of the examined fields appeared empty, with dark cell clusters at the edge of the empty areas. Moreover, the texture of the cells was irregular, as compared to controls.
Cultures morphology: Unfixed samples, stained by DAPI and propidium iodide (PI)As DAPI can pass through an intact cell membrane, it stains both live and dead cells; PI stains the nuclei after cells death, when the membrane does not prevent the passage of chemicals. The staining's showed a frame of cell death, increasing together with DZN concentration. Figure 1C and 1D show comparison between control slide and slide exposed to DZN 200ng/L stained with PI. The red dots show the presence of a scarce number of dead cells in controls, and a huge number of red nuclei in the clusters at the edge of the empty areas of the slide exposed to 200ng/L.
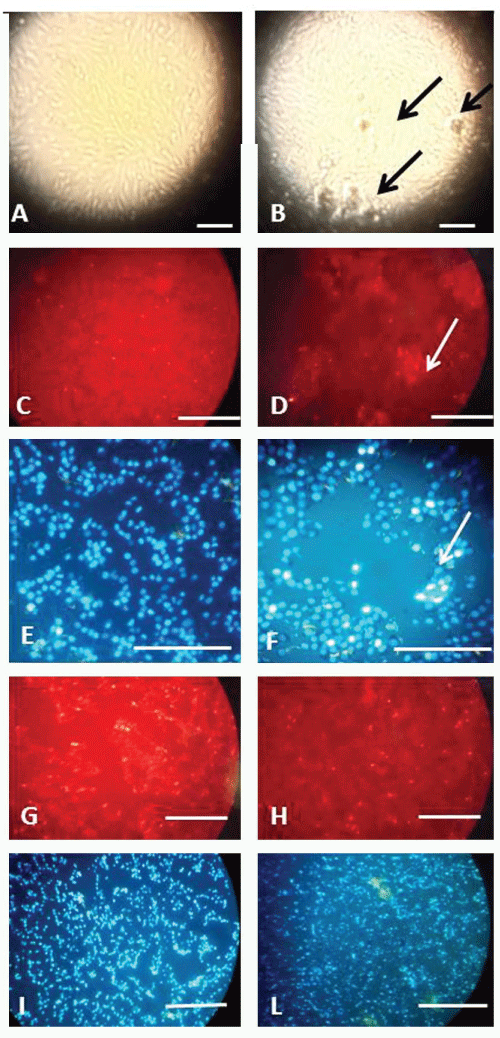
.
Figure 1: Morphology of C6 cell cultures exposed to DZN at different concentrations in culture medium: the dead cells are revealed by red dots (PI staining). The total population of cells is stained by white-blue fluorescence (DAPI staining).
A, C, E) control slides:
B , D, F) cultures exposed to 200ng/L.
G, I) cell cultures exposed to 0.2ng/L;
H, L) Cell cultures exposed to 0.002ng/L
Arrows point to clusters of dead cells surrounding empty areas. Bars equal 200 μm.
View Figure 1
DAPI staining showed the aspect of interphase nuclei of the control cells (Figure 1E) and confirmed the presence of the empty areas and picnotic nuclei characterized by very condensed and small nuclear material in the slide exposed to 200ng/L (Figure 1F).
The PI staining of slides exposed to 0.2ng/L showed the presence of small empty areas surrounded by apparently "unhealthy" nuclei in the cells, grouped to form islands scattered on the slide(Figure 1G). No empty areas in the slides exposed to 0.002ng/L (Figure 1H) were observed. The DAPI staining of slides exposed to 0.2ng/L confirmed the non-confluence of the cells (Figure 1I) While the cells exposed to 0.002ng/L presented fields full of cells (Figure1L).No apoptotic figures were seen either in controls or in exposed samples by both the stainings, suggesting a non-programmed cell death in the slides exposed to DZN, confirmed by the anomalous necrotic and picnotic aspect of the nuclei stained with PI and DAPI.
Occurrence of micronuclei (MN)
Figure 2 shows examples of the aspect of cells containing MNs, while the corresponding values of micronuclei presence are shown in figure 3.
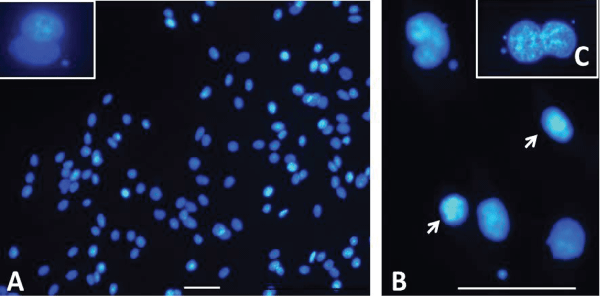
.
Figure 2: Cultured C6 cells, DAPI staining. Aspect of micronuclei in the control and exposed C6 cells.
A: Control, general view of part of a microscope field, B: Different aspects of cells exposed to DZN 200ng/L.
These presented nuclei and MN of different dimensions, and some picnotic (arrowheads) or swollen nuclei,
C: a late anaphase cell with 3 small MN. Bar equals 50 μm in all the images.
View Figure 2
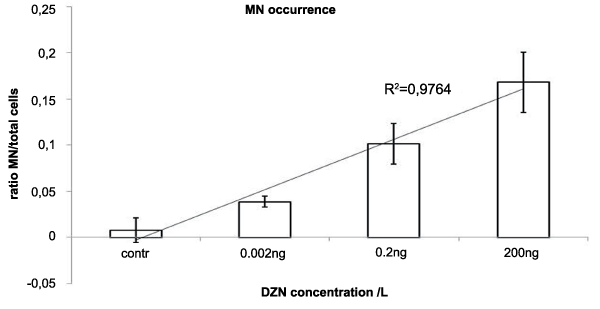
.
Figure 3: Occurrence of micronuclei in C6 glioma cells exposed to DZN. The X axis represents the concentrations of
DZN, the Y axis shows the ratio between the numbers of MNs/total number of cells.
View Figure 3
In the slides carrying control cells, only one field/six was found to contain 1MN (Figure 2A), while the exposed samples showed a major number of fields presenting cells with MNs in the cytoplasm (Figure 2B). In some cells more than 1MN was released in the same cell during cleavage (e.g.Figure 2C). Most MNs were found inside cytoplasm of anaphase cells (e.g. Figure 2C).
The number of micronuclei vs total cells in controls and exposed samples is reported in figure 3, followed by table 1 which shows statistical significance of data. For the statistical analysis, we counted all the micronuclei/field, independently from the occurrence in the same cell.
![]()
Table 1: Occurrenceof micronuclei
P values, One-Way Anova. (PAST program) Significant difference from controls is exhibited by the cells exposed to all the concentrations. ng: ng/L.
View Table 1
The slides exposed to the different concentrations of DZN showed a trend to increase MNs occurrence (Figure 3), The occurrence of MNs caused by the different exposures was significantly different among concentrations (Table 1), dose-dependence was confirmed by the high R-squared value of the trend line (R2 = 0.9764). This outcome shows that the MN occurrence is the most sensitive parameter among the others analyzed here.
Cholinesterase activities
IC50: Diazinon is not the most powerful ChE inhibitor among OP inscticides, as shown in figure 4. The IC50 values measured for purified E.e and rat brain AChE, is about 0.0456g/L, i.e. 45mg/L (far higher than the average of doses used in this work).In addition, the IC50 is different according to the AChE source, as shown by the the IC 50 for Electric eel AChE, lower than the one for rat brain AChE.The inhibition power seems to be very high for plasma BChE.
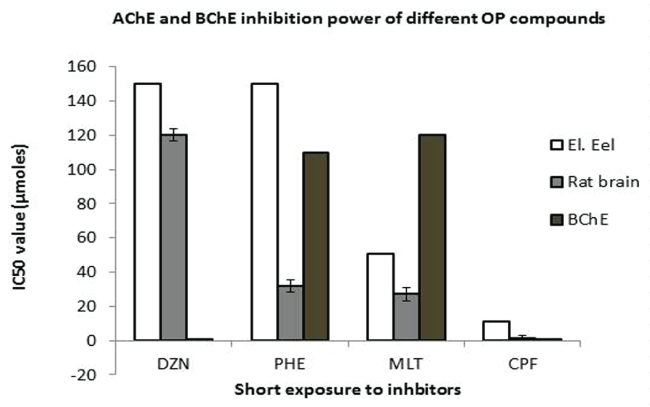
.
Figure 4: IC50 value expressed in μmoles. Inhibitors solved in MeOH (1mg/ml), final concentration of the MeOH in the incubation mixture was 0,5%, IC50 values are reported as means + S.D. of three independent experiments. Preincubation with inhibitor was 20 min, incubation time 20 min at 37°C rat brain vs electric eel P < 0,0001. El. eel: Electric eel, BChE: Mammalian Plasma BChE.
View Figure 4
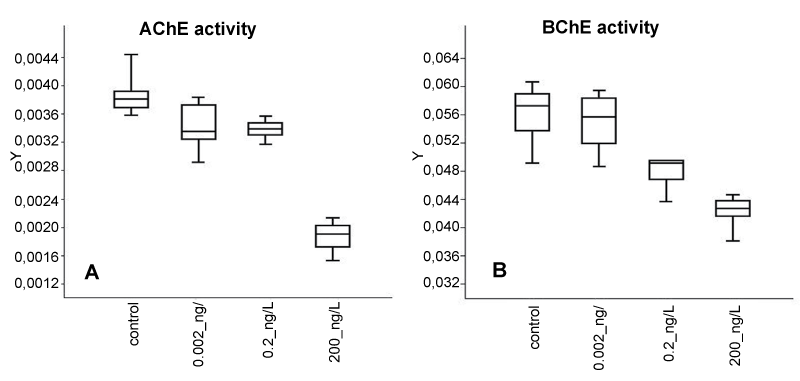
Low concentrations: The inhibition exerted by the exposures to Low dose DZN was not significantly different among the selected concentrations.For AChE significant difference was present only for control samples (different from all the other concentrations) and for the higher concentration (200ng/LM). For BChE activity, no significant inhibition change was present between control samples and the ones exposed to 0.002ng/L (Figure 5A, Table 2).
![]()
Table 2: Cholinesterse activities
Comparison of P.values for: A = AChE; B = BChE activities measured for different exposures. P values (One-Way Anova, PAST program).
View Table 2
.
Figure 5: Activity of AChE (E.C,. 3.1.1.7) and BChE (E.C. 3.1.1.8) X-axis = DZN concentration, A: Y-axis=AChE Units. B: Y-axis = μmoles of purified BTChI (Sigma) hydrolyzed/min/mg protein. AChE Unit = 1 μmole of ATChI hydrolyzed/min /mg protein.
View Figure 5
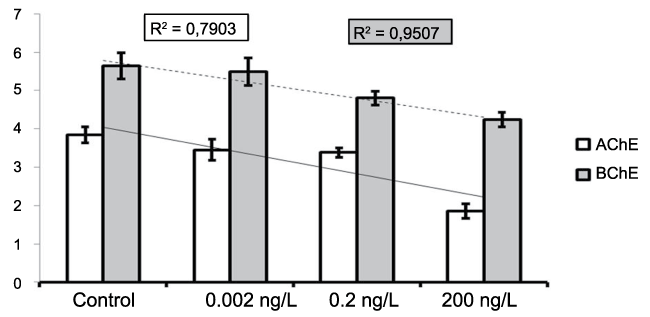
In figure 6, the dose-dependence is examined: For BChE, although the table 2 showsnon-significant difference in enzyme activity caused by the exposures to to the lower doses (0.002 and 0.2ng/L) a linear trend was present to decreasing, BChE the (=0.95) showed high fitness of the trend line to the data (Figure 6), thus suggesting dose-dependence. On the contrary, the squared-R (=0.79) of the trend linefor AChE inhibition shows scarce dose dependence. Thus, we consider that an effective inhibition on AChE is exerted only by the exposure to DZN 200 ng/L. Figure 6 also shows that BChE activity in C6 cells is generally higher than AChE, both in control and exposed samples.
.
Figure 6: Comparison between the ChE activities in control and exposed samples. X axis=DZN concentratios; Y axis=ChE Units (x103).
View Figure 6
Cell morphology and immunocytochemical reactions
In figure 7, the microscope objective was pointed on the slide portions outside the empty areas, in order to show the cell morphology and the immunoreactivity (IR) localization. The morphology of cells and localization of immunocytochemical reactions (IR) were affected by the exposure to the major DZN concentration (200ng/L), respectively, while the cells exposed to 0.002ng/L and 0.2ng/L showed aspects similar to the control ones (not shown).
.
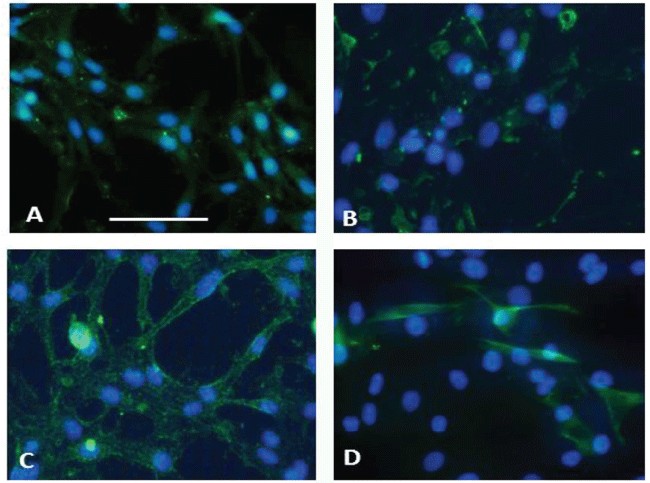
Figure 7: A: fibronectin-like IR expression in the C6 control cells; B: after exposure to 200ng/L DZN, the distribution is irregular, in dots localized at the surface of cells with swollen nuclei. C: ChAT-like IR in control cells is distributed at the cell surface; D: the cells exposed to DZN are distributed on different layers, and only the cells in the upper layer present IR in the cytoplasm and at the surface. Nuclei are counterstained by DAPI. Bars equal 50 μm. In all the cases the distribution of IR was different from the controls.
View Figure 7
The morphology of control cells was similar to the one reported by [22]. The control cells appeared spindle-shaped, with central nucleus and elongations at the two sides, very similar to the preadipocyte addressed to neurogenesis [23] (Figure 7A and Figure 7C). The cells exposed to DZN presented smaller cytoplasms and shorter elongations, swollen nuclei and different orientationas compared to the controls,
Localization of FN-like and ChAT-like immunoreactivity (IR)
FN-like IR is present in perinuclear localization and in granules present inside cytoplasm elongations of control cells (Figure 7A), while in samples exposed to 200ng/L DZN it is irregularly distributed in the cytoplasmic elongations of cells: the positive granules present in the cytoplasms are tiny; the nuclei are swollen if compared to controls (Figure 7 B). This may suggest that no new synthesis of FN or its precursors occurs in the exposed cells.
ChAT-like IR is widely distributed in the cytoplasm and at the surface of control cells (Figure 7C) as well as in cells exposed to0.2ng/L, not shown); the exposed cells are distributed on different layers (Figure 7D), and only some cells in the upper layer present IR in the cytoplasm and at the surface of . This suggests that new ACh synthesis is impaired by the exposure.
Discussion
Our outcomes are consistent with the concept that diazinon exerts stress effects on glial cells induction on gliotypic C6 cells. The effects can be observed since the lower concentrations [10,23,24], but seem independent from cholinesterase inhibition [10]. Indeed, in the present work this is suggested by the fact that the morpho-functional parameters tested in the present work are affected in a dose-dependent way, since the lower concentrations, while the inhibition of AChE and BChE activities seems less strictly linked to the doses. Actually, the effects on cell morphology and micronuclei occurrence match completely with the results of ChE inhibition only in cells exposed to the higher concentration (200 ng/L). This may be explained in the light of the fact that generally a number of enzymes represent secondary targets of OP inhibition [2]. The sum of these inhibitions may generate stress, in our case shown by the alteration of morpho-functional parameters. In addition, the micronuclei occurrence shows that cell loss is followed by very fast proliferation, as it occurswithin 24 h of treatment, while whole organisms need a longer exposure in order to exhibit similar aspects [18,25].
The second observation allowed by the present outcomes as compared with the current literature is that DZN toxicity is exerted in C6 cells with modalities only partially overlapping with the effects presented in other neurogenic cell types [11,26,27] e.g. as it concerns apoptosis. This is regulated and controlled by AChE [28,29] and this role is played in astrocytes, which are protected by acetylcholine against oxidative stress-induced apoptosis [30]. In the neurogenic NT2 cells, low doses (such as 0.3ng/L) of DZN caused apoptosis decrease in favor of cell viability [29] while in C6 cells viability was not enhanced, as demonstrated by loss of confluence and nuclear shape suggesting necrotic nuclear decay. Such a difference between different models agrees with the statement that DZN exerts different effects among the cell types [10,11] and shows that C6 cells are more sensitive to DZN than other cell types, in agreement with the finding [10] that DZN and other AChE inhibitors exert effect on DNA synthesis with preference toward C6 as compared with different neurogenic cells.
A third hypothesis suggested by the present outcomes is that the effects of the higher dose (200ng/L) may be related to AChE inhibition, and are probably due to the multiple roles that AChE exerts at cellular level [31]. For instance, we found that DZN, at concentrations up to 10 μM (i.e. 304mg/L) is not effective in inhibiting ChAT activity (data not shown). Thus, the response found in this work should be due to ChE inhibition, through cell-mediated events. The scarce number of cells expressing ChAT-like IR in samples exposed to 200ng/L as compared to controls may be due to the homeostatic no-need of new ACh synthesis. AChE inhibition causes a decreased ability to cleave acetylcholine and remove it from the receptorial sites [32,33]. Thus our data are consistent with the hypothesis proposed by other authors that the responsiveness of brain cells to stressors is a function of the level of ongoing cholinergic activity, in our case starting from DZN concentrations such as 200ng/L (the LOEL for effect on ChE and consequently on ChAT) and of the regulation of ChAT through a negative feedback mechanism [32,34]. In this light, fibronectin immunoreactivity (IR) also shows correspondence with AChE variation in samples exposed to DZN 200ng/L. The reduced localization of these molecules is not surprising. Actually, AChE molecules have been found strictly correlated to fibronectin during cell migration [35,36] and during axon elongation [37]. This suggests that DZN may exert an inhibiting effect on neuritogenesis, in partdue to the damage of DZN exposure on glial cells.
It is known that glial cells are actively involved in brain function, by exerting a number of functions [38,39], thus, collectively, these findings indicate that the impact of DZN on neurogenesismay act through different targets: 1- the brain-blood barrier dismantling due to glial cell death; 2- impairing functions of glial cells for expression of neurotrophic factors [24] and of molecules for nerve cells defense and functioning [37]; 3- reducing the synthesis of acetylcholine by impairing ChAT expression, 4- reducing cytoplasmic elongations by scarce production of fibronectin, the substrate supplied by glia to enhance neurite outgrowth [40].
References
-
Eggen T, Heimstad , Stuanes AO, Norli HR (2013) Uptake and translocation of organophosphates and other emerging contaminants in food and forage crops. Environ Sci Pollut Res Int 20: 4520-4531.
-
Casida JE, Quistad GB (2014) Organophosphate toxicology: safety aspects of nonacetylcholinesterase secondary targets. Chem Res Toxicol 17: 983-998.
-
Munro N (1994) Toxicity of the Organophosphate Chemical Warfare Agents GA, GB, and VX: Implications for Public Protection. Environ Health Perspect 102: 18-37.
-
Young JG, Eskenazi B, Gladstone EA, Bradman A, Pedersen L, et al. (2005) Association Between In Utero Organophosphate Pesticide Exposure and Abnormal Reflexes in Neonates. Neuro Toxicology 26: 199-209.
-
I.R.E.D Facts Diazinon; U.S. Environmental Protection Agency, Office of Prevention, Pesticides and Toxic Substances, Office of Pesticide Programs.
-
NPIC, National Pesticide Informantion Center, USA. Diazinon technical fact sheet.
-
Huen K, Bradman A, Harley K, Yousefi P, Boyd Barr D, et al. (2012) Organophosphate pesticide levels in blood and urine of women and newborns living in an agricultural community. Environ Res 117: 8-16.
-
Lu C, Toepel K, Irish R, Fenske RA, Barr DB, et al. (2006) Organic diets significantly lower children's dietary exposure to organophosphorus pesticides. Environ Health Perspect 114: 260-263.
-
Griffith W, Curl CL, Fenske RA, Lu CA, Vigoren EM, et al. (2011) Organophosphate pesticide metabolite levels in pre-school variability in a longitudinal study. Environ Res 111: 751-756.
-
Qiao D, Frederic J. Seidler, and Theodore A. Slotkin (2001) Developmental Neurotoxicity of Chlorpyrifos Modeled in Vitro: Comparative Effects of Metabolites and Other Cholinesterase Inhibitors on DNA Synthesis in PC12 and C6 Cells. Environmental Health Perspectives 109: 909-913.
-
Pope CN, Chakraborti TK, Chapman ML, Farrar JD, Arthun D (1991) Comparison of in vivo cholinesterase inhibition in neonatal and adult rats by three organophosphorothioate insecticides. Toxicology 68: 51-61.
-
Abbott NJ, Rönnbäck L, Hansson E (2006) Astrocyte-endothelial interactions at the blood-brain barrier. Nat Rev Neurosci. 7: 41-53.
-
Karpel R, Sternfeld M, Ginzberg D, Guhl E, Graessmann A, et al. (1996) Overexpression of alternative human acetylcholinesterase forms modulates process extensions in cultured glioma cells. J Neurochem 66: 114-123.
-
Ozawa A, Kadowaki E, Haga Y, Sekiguchi H, Hemmi N, et al. (2013) Acetylcholine esterase is a regulator of GFAP expression and a target of dichlorvos in astrocytic differentiation of rat glioma C6 cells. Brain Res 1537: 37-45
-
Bélanger M, Allaman I, Magistretti PJ (2011) Brain energy metabolism: focus on astrocyte-neuron metabolic cooperation. Cell Metab 14: 724-738.
-
Kruger NJ (1994) The Bradford Method for Protein Quantitation. Basic Protein and Peptide Protocols. Methods in Molecular Biology™ 32: 9-15.
-
Schmid W (1975) The micronucleus test. Mutation Research 31: 9-15.
-
Pietrapiana D, Modena M, Guidetti P, Falugi C, Vacchi M (2002) Evaluating the genotoxic damage and hepatic tissue alterations in demersal fish species: a case study in the Ligurian Sea (NW-Mediterranean). Mar. Pollut. Bull 44: 238-243.
-
Ellman GL, Courtney DK, Andres V Jr, Featherstone RM (1961) A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem. Pharmacol. 7: 88-95.
-
Lennet EDA (1978) An improved mounting medium for immunofluorescence microscopy. Am J Clin Pathol 69: 647-648.
-
Harlow E, Lane D (2006) Mounting samples in gelvatol or mowiol. CSH Protoc 2006.
-
Adornetto A, Pagliara V, Renzo GD, Arcone R (2013) Polychlorinated biphenyls impair dibutyrylcAMP-induced astrocytic differentiation in rat C6 glial cell line. FEBS Open Bio 3: 459-466.
-
Aluigi MG, Coradeghini R, Guida C, Scanarotti C, Falugi C (2009) Pre-adipocytes commitment to neurogenesis 1: preliminary localisation of cholinergic molecules. Cell Biol Int 33: 594-601.
-
Slotkin TA, Frederic J Seidler, Fabio Fumagalli (2007) Exposure to Organophosphates Reduces the Expression of Neurotrophic Factors in Neonatal Rat Brain Regions: Similarities and Differences in the Effects of Chlorpyrifos and Diazinon on the Fibroblast Growth Factor Superfamily. Environ Health Perspect 115: 909-916.
-
Bolognesi C, Creus A, Ostrosky-Wegman P, Marcos R (2010) Micronuclei and pesticide exposure. Mutagenesis 26: 19-26.
-
Falugi C, Rakonczay Z, Thielecke H, Guida C, Aluigi MG (2011) Cholinergic Pesticides. In: Stoytcheva M, Pesticides - The Impacts of Pesticides Exposure. INTECH Open Access Publisher: 221-242
-
Aluigi MG, Guida C, Falugi C (2010) Apoptosis as a specific biomarker of diazinon toxicity in NTera2-D1 cells. Chem Biol Interact 187: 299-303.
-
Zhang X, Lu L, Liu S, Ye W, Wu J (2013) Acetylcholinesterase deficiency decreases apoptosis in dopaminergic neurons in the neurotoxin model of Parkinson's disease. Int J Biochem Cell Biol 45: 265-272.
-
Zhang XJ, Greenberg DS (2012) Acetylcholinesterase involvement in apoptosis. Front MolNeurosci 5:40.
-
Liu Y, Zeng X, Hui Y, Zhu C, Wu J, et al. (2015) Activation of α7 nicotinic acetylcholine receptors protects astrocytes against oxidative stress-induced apoptosis: implications for Parkinson's disease. Neuropharmacology 91: 87-96.
-
Soreq H, Seidman S (2001) Acetylcholinesterase-new roles for an old actor. Nat Rev Neurosci 2: 294-302.
-
Tayebati SK, Di Tullio MA, Amenta F (2004) Effect of treatment with the cholinesterase inhibitor rivastigmine on vesicular acetylcholine transporter and choline acetyltransferase in rat brain. Clin Exp Hypertens 26: 363-373.
-
PahudG, SalemN, van de Goor J, Medilanski J, Pellegrinelli N, et al. (1998) Study of subcellular localization of membrane-bound choline acetyltransferase in Drosophila central nervous system and its association with membranes. Eur J Neurosci 10: 1644-1653.
-
Schrier BI, Thompson EJ (1974) On the Role of Glial Cells in the Mammalian Nervous System. Uptake, excretion, and rimetabolism of putative neurotransmitters by cultured glial tumor cells. The Journal of Chemistry 249: 1769-1760.
-
Somani SM, Babu SR, Arneric SP, DubeSN (1991) Effect of cholinesterase inhibitor and exercise on ChAT and acetylcholinesterase activities in rat brain regions. Pharmacol Biochem Behav 39: 337-343.
-
Gambardella C, Ferrando S, Gatti AM, Cataldi E, Falugi C, et al. (2015) Morphofunctional and biochemical markers of stress in sea urchin life stages exposed to engineered nanoparticles. Environ Toxicol.
-
Pizzurro DM, Dao K, Costa LG (2014) Diazinon and diazoxon impair the ability of astrocytes to foster neurite outgrowth in primary hippocampal neurons. Toxicol Appl Pharmacol 274: 372-382.
-
Falugi C, Aluigi MG (2012) Early appearance and possible functions of non-neuromuscular cholinesterase activities. Front Mol Neurosci 5: 54-63.
-
Barres BA (1991) New Roles for Glia. The Journal of Neuroscience 11: 3665-3694.
-
Akers RM, Mosher DF, Lilien JE (1981) Promotion of retinal neurite outgrowth by substratum-bound fibronectin. Dev Biol 86: 179-188.
