

International Journal of Pathology and Clinical Research
Cancer Stem Cell Microenvironment in Canine Glioblastoma Development: Animal Model for Human Disease
George Stoica*
Department of Veterinary Pathobiology, College of Veterinary Medicine and Biomedical Sciences, College Station, USA.
*Corresponding author:
George Stoica, DVM, MSc., PhD, Department of Veterinary Pathobiology, College of Veterinary Medicine and Biomedical Sciences, College Station, TX, 77843-4467, USA, Tel: (979) 845-5089; E-mail: gstoica@cvm.tamu.edu
Int J Pathol Clin Res, IJPCR-1-014, (Volume 1, Issue 1), Review Article; ISSN: 2469-5807
Received: September 19, 2015 | Accepted: November 25, 2015 | Published: November 30, 2015
Citation: Stoica G (2015) Cancer Stem Cell Microenvironment in Canine Glioblastoma Development: Animal Model for Human Disease. Int J Pathol Clin Res 1:014. 10.23937/2469-5807/1510014
Copyright: © 2015 Stoica G. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Abstract
Understanding the cancer stem cells (CSCs) role in malignancy is based on a comprehensive understanding of normal stem cells role and plasticity in maintaining the normal homeostasis or regeneration in various organ tissues. The niche microenvironment that sustains and regulates stem cells is tightly regulated by physical factors and hormonal signals. In the central nervous system (CNS) malignancy the CSCs niche is deregulated by intrinsic and extrinsic factors, which disrupt the homeostatic balance from quiescence state to proliferation and differentiation that characterized glioblastoma development. Here, we review the role of microenvironment in development of glioblastoma in the dog species as an animal model for human disease for a better understanding of underlined mechanism(s) and possible target therapy for the benefit of both species. A comparative approach to the study of canine glioblastoma provides an excellent animal model for human glioblastoma pathogenesis and development of novel therapies in both species.
Keywords
Cancer stem cells, Canine glioma, Microenvironment
Introduction
The cancer stem cell hypothesis in glioma implies that tumor development, like normal tissue development, relied on a unique subset of tumor cells (CSCs) with the ability to self-renew and multi-lineage differentiation which constitute the tumor mass. Recent evidence generated from studies in solid tumors demonstrated that CSCs reside in a niche. This hypothesis derives from reports on normal stem cells from various tissues, showing that stem cells exist within protective niches that are composed of a number of differentiated cell types [1,2]. This cellular microenvironment provides direct cell contacts and secreted factors that maintain stem cells in a quiescent state. Histological observations of rodent tissues suggest that neural stem cells lie within a vascular niche in which endothelial cells regulate stem cell self-renewal. In glioblastoma (GB) it is hypothesized that vascular endothelial cells provide such a niche for the CSCs similar to the situation with normal neuronal stem cells [3].
The study of tumorigenesis and the evaluation of new therapies for GB require accurate and reproducible brain tumor animal models, which ideally should recapitulate key features of the human disease, be reproducible, and resemble progression kinetics and anti-tumor immune responses of spontaneous GB. Human glioma xenografts implanted in immuno compromised mice are extensively used. However, their xenogeneic nature impairs the study of immune-mediated anti-tumor strategies. Syngeneic murine models, such as GL26 mouse glioma cells in C57BL6 mice [4,5] and CNS1 rat glioma cells in Lewis rats [6] are non-immunogenic. Thus, syngeneic glioma models would be relevant for studying the response of brain tumors to immunotherapy [7]. Molecular and cytogenetic characterization of canine primary brain tumors is advancing rapidly since the publication of the canine genome. Many similarities to human tumors have already been described, relating to chromosomal instability, expression profiles, the presence of stem-like tumor cells, expression of growth factors and their receptors such as EGF, PDGF, VEGF, as well as other markers frequently described in human tumors such as IL-13Rα2, IGFBP2 and telomerase activity [7]. Limited data are available relating to the efficacy of standard therapeutic modalities in canine glioblastomas and it is likely that some significant differences will be present between human and canine gliomas, as has been described for meningiomas [8-10].
In summary, the translational potential of rodent GB models to develop novel therapeutic approaches for human GB is still controversial. Spontaneous glioblastoma in dogs reproduce most typical human glioblastoma characteristics and constitute an excellent large animal model, which could help to better understand the pathogenesis and predict the effectiveness of novel therapeutic approaches in human trials.
Ideally, brain tumor models should be of glial origin, grow in vitro and in vivo with predictable and reproducible growth patterns, have close resemblance in vivo to the histopathology of human glioblastomas, be weakly or non-immunogenic and ultimately, their progression in vivo should be highly reproducible. Unfortunately, the ideal GB animal model that comprises all of the above features does not exist [10].
In veterinary medicine there is a considerable literature that describes the occurrence of spontaneous tumors of the central nervous system in animals [11,12]. Astrocytoma, of which glioblastoma multiforme (GBM), the most malignant form, is one of the most common neoplasms of the central nervous system in animals. Recent reports also demonstrate the similarity between human and dog GBM and emphasize the importance of studying dog spontaneous brain tumors in order to elucidate the mechanism(s) of tumor dispersal, post surgical reoccurrence and resistance to therapy [11,13-15].
Normal Stem Cell Homeostasis
Adult mammalian tissues are diverse in regard to their structure and functions that need continuously to adapt to survive and growth by maintaining a proper regulated homeostatic microenvironment. This microenvironment is regulated at the cellular level by the so call stem cells (SCs) that populate every tissue. However, turnover rate of SC is not the same in every tissue, some like skin, intestine have a greater rate of replication then liver, muscle or kidney with barely detectable rate of proliferation. Fallowing pathological conditions, SCs reverse rapidly to proliferating mode to regenerate the loss tissue and reverse to normal homeostatic environment in the tissue affected [16]. Tissue repair can be achieved also by dedifferentiation alone in the absence of a reserve stem cell population. Differentiation cells are able to generate cycling progenitors of a different lineage. For example, in the central nervous system (CNS) following vascular stroke in mouse brain, lineage tracing provides evidence for differentiated astrocytes generating proliferating neuroblasts, supplementing neuroblasts derived from mobilized neural SCs in the SVZ which have migrated into the zone of injury (Figure 1) [7,16].
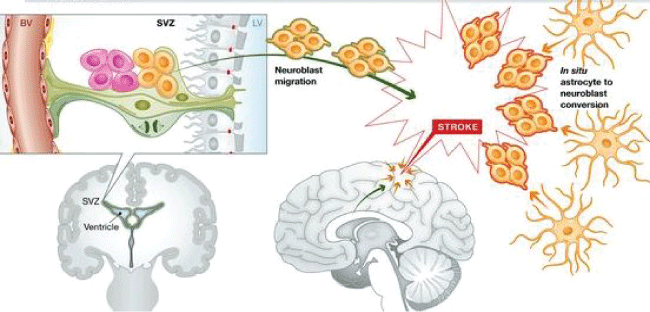
.
Figure 1: Neural SC responses to stroke. Following a CNS stroke, which results in cell death due to ischemia, clusters of neuroblasts (orange) appears in the injured region. Lineage tracing argues these derive both from mobilization of neural SCs from the SVZ followed by migration to the site of injury (orange with green border) and from differentiated astrocytes within the area of the stroke entering a neurogenic programme (orange with red borders) [16].
View Figure 1
Consequently, we defined the SCs as a cell population that maintain long-term tissue turnover and contribute to tissue repair after injury. The plasticity of SCs might extend to differentiating cells, which in pathological conditions are capable of reverting to stemness condition.
Significance of Niche in Maintaining Stem Cells
The SCs resides in a specific functional environment call niche, the microenvironment that regulates stem cells by physical factors and hormonal signals. In rodent and human brain adult neural SCs are found in the subventricular zone (SVZ) lining the lateral ventricles and in the dentate gyrus (DG) of the hippocampus in a quiescent state. SCs are capable of both asymmetric and symmetric divisions generating two SCs or two differentiating astroglial or neural progenitor daughters [17]. After division, some SCs return to a quiescent state, negative for proliferation markers, paralleling the transition between dormant and active states seen in hematopoietic SCs. Paracrine niche signals also regulate neural SCs in the SVZ, which extend cellular processes into the cerebrospinal fluid (CSF) that fills the ventricles and forms contacts with underlying endothelial cells [18,19]. These contacts allow regulation of neural SCs via both secreted factors in the CSF and signals from the vasculature. For example, endothelium-derived Ntf3 and G protein-coupled receptor ligands in the CSF promote SC quiescence [19,20].
The nervous system homeostatic microenvironment plays an essential role in systemic physiological SC regulation. This concept was demonstrated first in hematopoietic stem cell (HSC), in chronic physiological stress induces increased levels of leucocytes, as a proof of concept. This effect is mediated by noradrenaline release from sympathetic neurons in the bone marrow, which decreases production of Cxcl12 in niche cells triggering HSC proliferation [21]. Understanding how neural, endocrine and cytokine signals regulate cell production to meet the requirements of an organism is a key challenge for SC research.
In summary tissues with a low rate of turnover, in rodent and human brains, believed to be post-mitotic, continue to proliferate new neurons throughout life, which contribute to specific forms of learning and memory and responding to stress [22-25].
Niche and Cancer Stem Cells (CSc) in Glioma Development in Animal Models
There is a general acceptance in the field of oncology that CSCs in solid tumors reside in a niche. This hypothesis derives from reports on normal stem cells from various tissues, showing that stem cells exist within protective niches that are composed of a number of differentiated cell types [1,2]. This cellular microenvironment provides direct cell contacts and secreted factors that maintain stem cells in a quiescent state. Histological observations of rodent and monkey tissues suggest that neural stem cells lie within a vascular niche (angiogenic niche) in which endothelial cells regulate stem cell self-renewal. In gliomas it is hypothesized that vascular endothelial cells provide such a niche for the glioma CSCs similar to the situation with normal neuronal stem cells [3,16,26].
The cancer stem cell hypothesis implies that tumors contain a minority of cells with stem-like properties, which have the ability to self-renew and sustain the growth of the tumor [27]. Tumor stem cells were first identified in hematopoietic malignancies and more recently were also demonstrated in various solid tumors, including tumors of the brain such as glioblastomas, medulloblastomas and ependymomas [15,28-30]. Brain tumor stem cells in gliomas are defined by (i) the capacity to self-renew, (ii) the ability to initiate brain tumors upon orthotopic implantation and (iii) multipotency, that is, the capacity to differentiate into cells with a neuronal, astrocytic, or oligodendroglial phenotype [31,32]. However, multilineage differentiation is not a requirement of the CSC. It is more important that CSCs regenerate the cell types observed in the original tumor [33]. In addition, brain tumor stem cells are characterized by the expression of neural stem cell antigens and the ability to grow as nonadherent spheres termed neurospheres when cultured in the presence of epidermal growth factor (EGF) and fibroblast growth factor (FGF) under serum-free conditions. However, not all the neurospheres arise from stem cells, finding that negatively impacts on the idea that sphere forming assays are a functional unique assay for detecting in vivo stem cells. Sphere-forming assays evaluate the potential of a cell to behave as a stem cell when removed from its in vivo niche [34]. Neurospheres are composed of stem cells in small numbers, progenitors and differentiated cells constitute the majority. Consequently, neurosphere assay cannot be used alone to define the in vivo stem cells. Thus, stem-like cells in brain tumors share many characteristics with normal neural stem cells, supporting the hypothesis that brain tumors can arise from neural stem or progenitor cells [32]. Recent studies further show that glioblastoma cells cultured under neural stem cell conditions can display heterogeneous growth characteristics and molecular profiles, suggesting that they may either arise from different cell types or from similar cells that have acquired different genetic alterations [35]. Whether CSCs arise from normal stem cells, progenitor cells, or differentiated cells is still a contentious issue at the present time. Nevertheless, CSCs in gliomas express CD133 and nestin that mark neural and progenitor cells [28]. The factors governing glioblastoma cell differentiation and migratory potential and therefore tumor phenotype, are not completely understood. On the basis of gene expression profiling and by neurobiological criteria, two subtypes were identified recently among human glioblastoma cultures established under neural stem cell conditions. One cluster of CSCs expressed neurodevelopmental genes and displayed a full stem-like phenotype. The second cluster displayed an expression signature for extracellular matrix- (ECM)-related genes and only a restricted stem-like phenotype, fulfilling only in part the criteria considered typical of glioblastoma stem cells [36]. Whether these differences reflect variations in stem-like cells present between individual glioblastomas and/or distinct emerging lineages remains to be established.
Significance of Homeostatic Environment in Dog Glioblastoma Development
Increased life expectancy of companion animals and improvements in veterinary medical practice has led to an increase incidence of cancer, one of the main causes of death in dogs and cats [13,14]. Of the domesticated animal species, most examples are seen in dogs, with much lower occurrence in other species. Intracranial neoplasia occurs more frequently in dogs than in humans [37].
Canine intracranial glioblastoma occurs most commonly in brachycephalic breeds, in particular the Boxer and Boston terrier. No sex predilection has been reported and they are recognized with greater incidence in animals over 6 years of age. Clinical signs caused by gliomas may vary with tumor location and neurological deficits reflect direct nervous tissue involvement as well as secondary effects of peritumor edema, necrosis, hemorrhage, compression, herniation, or obstructive hydrocephalus. Cerebrospinal fluid findings in animals with astrocytoma are usually non-specific and include increased protein concentration and mixed-cell pleocytosis that reflect disturbance of the blood-brain-barrier. Computed tomography (CT) and magnetic resonance imaging are routinely available to veterinary practitioners for diagnosis and localization of intracranial lesions. Computed tomographic characteristics of some canine brain tumors have been reported to be similar to those in humans [38,39].
Glioblastoma multiforme (GBM), grade IV [40] is the most common and lethal primary malignant brain tumor in human and dog. The highest incidence (30%) of this tumor occurs in the Boxer [41]. Despite progress in research on the molecular aspects of GBMs, the prognosis of these brain tumors continues to be dismal in humans and animal species. One reason for the lack of clinical advances is ignorance of the cellular origin of this disease and lack of understanding of the mechanisms of glioma cell migration and dispersal, which delays the application of molecular analyses to treatment and impairs anticipation of tumor biological behavior. The CSCs as the cell of origin and their contribution in glioblastoma development and tumor invasion is complex and controversial. In a recent seminal review article, Wabik et al. mentioned the role in tissue repair of SCs in the mouse brain following vascular stroke when astrocytes mobilized at the site of injury and generate proliferating neuroblasts [16]. This finding implies that astrocytes and other neural cells in specific pathological conditions can dedifferentiate and participate in tissue repair and potentially in CNS tumor development. In case of brain injury of any etiology the SCs participating in tissue repair process can accumulate genetic and epigenetic mutations, which could favor glioblastoma development. In animal and human CNS tumors can develop in various anatomic locations some at distant from SVZ, the site of SCs origin. This implies that cells of origin for glioblastoma can have different origin raising the hypothesis involving other neurogenic cells, which under specific conditions, can reverse to the stemness status and participate in tumor development.
Glioblastoma is a nonmetastatic tumor, but highly locally invasive, diffusely disseminating into the brain parenchyma and possessing cancerous cells outside the margin of therapeutic intervention [40-43]. The GBM infiltrative path into the normal brain is not random; it often follows white matter tracts and extends along perivascular spaces, the glial limitans externa and the subependyma. Their diffuse infiltration suggests the activation of genetic and cellular programs that distinguish them from cells in the tumor core. Additionally, migrating tumor cells may activate the host brain microenvironment to facilitate tumor dispersal.
A recent report demonstrated that GBM isolated from a Boxer dog (D-GBM) has cells with phenotypical characteristics of CSCs. The intracranial tumor was located within the brain lateral ventricle, anatomical location, which intuitively implied that the CSCs in this case originated from SVZ zone. As a proof of concept, this cell line's morphophenotype expressed cell markers such as nestin and CD133. CSCs demonstrated glial and neuronal differentiation when exposed to differentiation growth factors in vitro. An intracranial orthotopic model using nude mice was utilized for in vivo evaluation of D-GBM tumorigenicity. Immunohistochemistry results of nude mouse xenografts also demonstrated glial and neuronal differentiation. These data also demonstrate the similarity between human and dog GBM and emphasize the importance of studying dog spontaneous brain tumors in order to elucidate the mechanism(s) of tumor dispersal, post surgical reoccurrence and resistance to therapy [15].
Glioblastoma invasion is a combination of the ability of CSCs to migrate and their ability to modulate the extracellular matrix (ECM). The ECM of the brain is distinct from the ECM of most organs, being ill defined and scant. The matrix consists primarily of hyaluronic acid, except for the areas around the vessels and the pial surface (glial limitans) where there is well-defined basal lamina that include collagen. Glioma cell invasion preferentially involves the perivascular and subpial spaces, where basal lamina is well-defined, but also involves perineuronal and white matter locations in which the ECM is ill-defined.
Cancer stem cell migration in gliomas is a multifactorial event regulated by the interrelation between a minority of tumor cells with stem cell-like properties and the host microenvironment. Cellular heterogeneity, necrosis and neovascularization constitute the morphological phenotype that characterizes gliomas. Necrosis and angiogenesis in gliomas, occurs in a hypoxic environment. A working hypothesis that links tumor hypoxia, necrosis and angiogenesis appears to emerge in glioblastoma.
GBM is one of the most lethal primary brain tumors in human and animal species. This tumor is comprised of a phenotypically heterogeneous cell population [15,41,42]. The mechanisms responsible for the GBM heterogeneity and the ability of tumor cells to migrate into the brain parenchyma are incompletely understood. Localized hypoxia appears to upregulate migration-associated genes, leading to migration of tumor cells away from a central hypoxic center. Necrosis is a central feature of the highest-grade malignant gliomas. No histological feature is more powerful in predicting poor prognosis [40,42]. There is a vicious cycle involving necrosis-induced hypoxia and various angiogenic and growth factors that combine to foster the highly malignant state of glioblastoma. In the setting of rapidly dividing cells with high metabolic demands, small regions of necrosis may develop in areas where metabolic demands exceed supply. Another recently suggested possibility is that in response to hypoxia, small clones of tumor cells acquire molecular characteristics that encourage more active migration, thus clearing a central region more susceptible to necrosis. This hypothesis is based on work showing that glioblastoma tumor cells surrounding necrotic centers (pseudopalisades) are less proliferative and more apoptotic than adjacent cells [43]. These perinecrotic cells also express hypoxia-inducible genes, such as hypoxia-inducible factor 1 alpha (HIF-1α) and in vitro studies have demonstrated that hypoxia increases cellular migration and gelatinase activity [44,45]. Hypoxic cell death may lead to the release of growth factors from the dying cells and some growth factors may remain intact within the necrotic zones. More importantly, hypoxia may allow the emergence of resistant and thus high malignant clones of tumor cells. When selected molecules that are preferentially expressed in palisading cells are used as prognostic markers in glioblastomas, they correlate with shorter patient survival [46]. This supports the notion that these perinecrotic regions are zones in which more malignant clones are actively selected. In addition, the interactions between hypoxia, necrosis, upregulation of growth factors and selection of malignant clones might provide an explanation for the marked histological heterogeneity noted in glioblastomas and a possible explanation for the marked resistance to conventional cytotoxic therapies seen in clinical patients with glioblastoma (Figure 2) [40]. Clonal evolution and CSCs models are not mutually exclusive but rather co-exist and complement each other in glioblastoma [14].
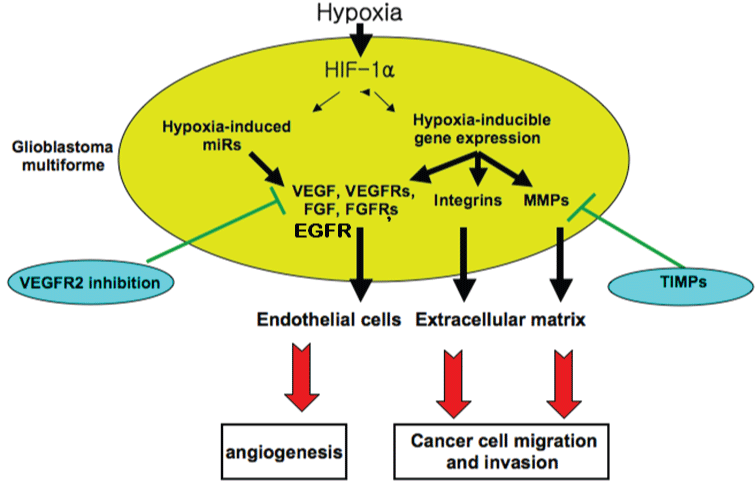
.
Figure 2: Schematic representation of the role of hypoxia and hypoxia-induced factors in glioblastoma development and potential tumorigenic inhibitors.
View Figure 2
The main challenge presented by gliomas is the tendency of the malignant cells to invade the adjacent normal brain tissue. Extracellular matrix (ECM) degrading proteases are extremely important in mediating gliomas invasiveness [47,48]. The cellular heterogeneity of gliomas indicates that they use different cell-surface receptors for their migration, probably depending on the available cell substrate. Several ECM components and their receptors have been shown to have an important role in glioma-cell migration and invasion. Serine, cysteine and metallo- proteinases (MMP) have important roles with regard to brain tumors, as well as other types of cancer. The remarkable similarities between these proteases include the fact that their level of expression is significantly increased in most cancers, they are present at the leading edge of the tumour and they are prognostically significant during tumour progression. Some evidence exists for the coordination of proteolysis across protease families. Proteolytic cleavage of adherent ECM proteins can indirectly stimulate cells to produce other proteases through changes in the interaction (affinity or avidity) of integrins with the substrate after cleavage. For example, serine protease urokinase-type plasminogen activator (uPA) has been shown to increase MMP activity indirectly through the interactions of its receptor uPAR with integrin receptors [47,49]. Furthermore, interactions between integrins expressed by glioma cells and the ECM and the activity of MMPs, form the basis for glioma-cell migration and invasion [50]. As such, the net proteolytic and therefore invasive, potential of a given tumour cell might depend on the interplay between many enzymes [47].
Tumor progression leading to metastasis or invasion appears to involve equipping cancer cells with the appropriate adhesive (integrin) phenotype for interaction with the ECM. The integrins constitute a family of transmembrane receptor proteins composed of heterodimeric complexes of noncovalently linked alpha and beta chains. Integrins function in cell-to-cell and cell-to-ECM adhesive interactions and transduce signals from the ECM to the cell interior and vice versa. Previous studies reported the expression of various integrins in brain tumors [51,52]. There have been few investigations of the expression of αvβ5 and αvβ3 in gliomas. Few data are available regarding the relationship between the expression of these integrins angiogenesis and invasion in this type of tumor. Two αv integrins, αvβ3 and αvβ5, are necessary for cytokine- or tumor-induced angiogenesis. They mediate two distinct pathways of angiogenesis expression of αvβ3 is required for fibroblast growth factor 2 (FGF-2) and tumor necrosis factor α-induced angiogenesis. The integrin αvβ5 is required for VEGF- and transforming growth factor β-induced angiogenesis [53]. Hypoxia has been reported to stimulate expression of integrins αvβ3 and αvβ5 in retinal microvascular endothelial cells and to inhibit expression of integrin β1. The hypoxic induction of the αv integrin is partially mediated through VEGF induction in an autocrine-paracrine manner [54]. The angiogenic integrins have been reported to not only mediate vascular cell migration but also to regulate metalloproteinase activity and cell proliferation. These specific increases in the tumor cellular integrin expression may contribute to neovascularization and invasion observed in dog glioblastomas. The results from our lab of integrins expression are illustrated in figure 3 [15].
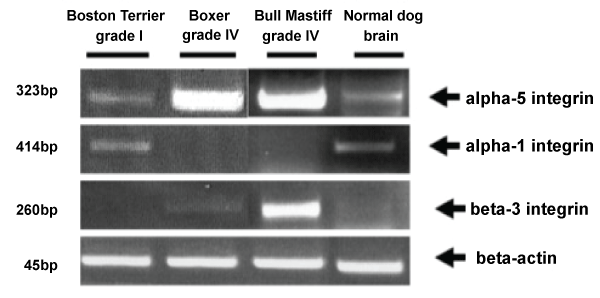
.
Figure 3: Integrins comparative expression in normal brain and different grades of glioblastoma in dog. The RT-PCR analysis of integrins was performed in dog glioblastoma multiforme (D-GBM, grade IV) cells isolated from a Boxer, a pilocytic astrocytoma (PCA, grade I, WHO classification, 2007, 42), primary cell culture isolated from a Boston terrier, a grade IV GBM cells isolated from a Bull Master dog and normal primary culture from a Boxer dog brain. Our results demonstrated that grade IV GBM expresses higher level of α-5 integrin compared with grade I PCA tumor. α-1 integrin was not detected in grade IV GBM and normal primary culture from a Boxer dog. Beta-3 integrin expression was higher in grade IV DBM compared with grade I PCA.
View Figure 3
The intracranial brain tumors developed in a rich neuroendocrine stimulatory/inhibitory homeostatic environment that could participate directly or indirectly in the development and migratory potential of glioblastomas. Recent reports begin to recognize the role of neurotransmitters in regulating tumor phenotypes. A recent Science article reported the potential role of parasympathetic cholinergic fibers in prostate cancer tumor dissemination [55].
A recent report highlights the importance of the nerve elements in the tumor microenvironment [56]. This report underlines that neurotransmitters, such as dopamine, are required for glioblastoma growth and provides one explanation for its unusual phenotype. An intriguing issue in glioblastoma therapeutic development involved the observation that while Epidermal Growth Factor Receptor (EGFR) deregulation is critically important in the pathogenesis of glioblastomas [57], EGFR inhibitors are clinically ineffective [58]. The cross-signaling between dopamine receptor DRD2 and EGFR offers an explanation for the poor clinical efficacy of inhibitors of receptor tyrosine kinases such as EGFR. It is suggested that during glioblastoma pathogenesis, EGFR re-wired the molecular circuitry of the astrocyte as to "hijack" the neurotransmitter-mitogenic signaling axis [59]. Consequently, the high concentrations of neurotransmitter in the CNS signal to effector molecules downstream of EGFR, thereby bypassing the need for EGFR activation.
Considering dopamine roles in reward systems in the brain, emotion and personality traits, raises the possibility that personality and emotion may impact the risk of glioma development or growth through modulation of dopamine release. Interestingly, studies have related cancer risks to personality types [56,60]. Therapeutic synergy between an EGFR inhibitor and a DRD2 antagonist, haloperidol, against glioblastomas using independent models have suggested that patients undergoing antipsychotic therapy are less likely to suffer from brain cancer [61]. It is hypothesize that non-CNS microenvironment may not possess the level of neurotransmitters required to sustain glioblastoma growth. In this context, drugs that modulate neurotransmitter function warrant consideration as antineoplastic agents in glioblastoma.
Conclusions
Multiple autocrine/paracrine signaling systems are involved in the development, maintenance and invasion of glioblastoma. Invasion of grade IV glioblastoma usually follows white matter tracts and extends along perivascular spaces, the glial limitans externa and the subependyma, which implicates scaffolding mechanical plasticity necessary for cell motility. This histological observation suggests that normal brain tissues, which surround the glioma, are significantly important in tumor development and progression. Consequently, invasive glioma cells receive paracrine signals from the proximity of normal cells encountered in their tracts, which support tumor invasion. Interaction of tumor cells with the peritumoral tissues could result in severe disruption of brain homeostasis.
The interconnection between CSC and homeostatic microenvironment is of significant clinical importance for better understanding of failing therapeutic approaches in glioblastoma treatment in human and dog species. Therapeutic modalities of targeting CSCs in glioblastoma might not be sufficient to eradicate the tumor or increase the survival time of affected patients. A combine therapeutic modality involving suppression of CSC development and annihilation of the negative brain microenvironment response to tumor invasion might be an optimal approach to glioblastoma treatment. Given the relative high incidence and similarity of glioblastoma in dog specie with human counterpart it might provide an excellent animal model for understanding the mechanism(s) of glioma and development of novel therapeutic paradigms.
Conflict of interest
The author of this review article has no financial or personal relationship with other people or organizations that could inappropriately influence or bias the content of the paper.
References
-
Fuchs E, Tumbar T, Guasch G (2004) Socializing with the neighbors: stem cells and their niche. Cell 116: 769-778.
-
Pierret C, Spears K, Morrison JA, Maruniak JA, Katz ML, et al. (2007) Elements of a neural stem cell niche derived from embryonic stem cells. Stem Cells Dev 16: 1017-1026.
-
Calabrese C, Poppleton H, Kocak M, Hogg TL, Fuller C, et al. (2007) A perivascular niche for brain tumor stem cells. Cancer Cell 11: 69-82.
-
Albright L, Madigan JC, Gaston MR, Houchens DP (1975) Therapy in an intracerebral murine glioma model, using Bacillus Calmette-Guerin, neuraminidase-treated tumor cells and 1-(2-chloroethyl)-3-cyclohexyl-1-nitrosourea. Cancer Res 35: 658-665.
-
Curtin J, King GD, Xiong W, Liu C, Lowenstein PR, et al. (2005) Stimulation of the immune system using gene therapy is an effective approach for treating brain tumors in mouse and rat models. American society of gene therapy 8th annual meeting. Mol Ther 11: S106
-
Kruse CA, Molleston MC, Parks EP, Schiltz PM, Kleinschmidt-DeMasters BK, et al. (1994) A rat glioma model, CNS-, with invasive characteristics similar to those of human gliomas: a comparison to 9L gliosarcoma. J Neurooncol 22: 191-200.
-
Magnusson JP, Goritz C, Tatarishvili J, Dias DO, Smith EM, et al. (2014) A latent neurogenic program in astrocytes regulated by Notch signaling in the mouse. Science 346: 237-241.
-
Dickinson PJ, LeCouteur RA, Higgins RJ, Bringas JR, Larson RF, et al. (2010) Canine spontaneous glioma: a translational model system for convection-enhanced delivery. Neuro Oncol 12: 928-940.
-
Thomas R, Duke SE, Wang HJ, Breen TE, Higgins RJ, et al. (2009) 'Putting our heads together': insights into genomic conservation between human and canine intracranial tumors. J Neurooncol 94: 333-349.
-
Candolfi M, Curtin JF, Nichols WS, Muhammad AG, King GD, et al. (2007) Intracranial glioblastoma models in preclinical neuro-oncology: neuropathological characterization and tumor progression. J Neurooncol 85: 133-148.
-
Stoica G, Kim HT, Hall DG, Coates JR (2004) Morphology, immunohistochemistry and genetic alterations in dog astrocytomas. Vet Pathol 41: 10-19.
-
Summers BA, Cummings JF Summers BA, Cummings JF, Alexander de Lahunta (1995) Tumor of the central nervous system. In: Veterinary Neuropathology, Mosby, St. Louis, MO, 351-395.
-
Pang LY, Argyle D (2010) Cancer stem cells and telomerase as potential biomarkers in veterinary oncology. Vet J 185: 15-22.
-
Pang LY, Argyle DJ2 (2015) The evolving cancer stem cell paradigm: implications in veterinary oncology. Vet J 205: 154-160.
-
Stoica G, Lungu G, Martini-Stoica H, Waghela S, Levine J, et al. (2009) Identification of cancer stem cells in dog glioblastoma. Vet Pathol 46: 391-406.
-
Wabik A, Jones PH (2015) Switching roles: the functional plasticity of adult tissue stem cells. EMBO J 34: 1164-1179.
-
Bonaguidi MA, Wheeler MA, Shapiro JS, Stadel RP, Sun GJ, et al. (2011) In vivo clonal analysis reveals self-renewing and multipotent adult neural stem cell characteristics. Cell 145: 1142-1155.
-
Silva-Vargas V, Crouch EE, Doetsch F (2013) Adult neural stem cells and their niche: a dynamic duo during homeostasis, regeneration and aging. Curr Opin Neurobiol 23: 935-942.
-
Codega P, Silva-Vargas V, Paul A, Maldonado-Soto AR, Deleo AM, et al. (2014) Prospective identification and purification of quiescent adult neural stem cells from their in vivo niche. Neuron 82: 545-559.
-
Delgado AC, Ferron SR, Vicente D, Porlan E, Perez-Villalba A, et al. (2014) Endothelial NT-3 delivered by vasculature and CSF promotes quiescence of subependymal neural stem cells through nitric oxide induction. Neuron 83: 572-585.
-
Heidt T, Sager HB, Courties G, Dutta P, Iwamoto Y, et al. (2014) Chronic variable stress activates hematopoietic stem cells. Nat Med 20: 754-758.
-
Aimone JB, Li Y, Lee SW, Clemenson GD, Deng W, et al. (2014) Regulation and function of adult neurogenesis: from genes to cognition. Physiol Rev 94: 991-1026.
-
Ernst A, Alkass K, Bernard S, Salehpour M, Perl S, et al. (2014) Neurogenesis in the striatum of the adult human brain. Cell 156: 1072-1083.
-
Snyder JS, Soumier A, Brewer M, Pickel J, Cameron HA (2011) Adult hippocampal neurogenesis buffers stress responses and depressive behaviour. Nature 476: 458-461.
-
Spalding KL, Bergmann O, Alkass K, Bernard S, Salehpour M, et al. (2013) Dynamics of hippocampal neurogenesis in adult humans. Cell 153: 1219-1227.
-
Walton RM (2012) Postnatal neurogenesis: of mice, men and macaques. Vet Pathol 49: 155-165.
-
Clarke MF, Fuller M (2006) Stem cells and cancer: two faces of eve. Cell 124: 1111-1115.
-
Galli R, Binda E, Orfanelli U, Cipelletti B, Gritti A, et al. (2004) Isolation and characterization of tumorigenic, stem-like neural precursors from human glioblastoma. Cancer Res 64: 7011-7021.
-
Lee J, Kotliarova S, Kotliarov Y, Li A, Su Q, et al. (2006) Tumor stem cells derived from glioblastomas cultured in bFGF and EGF more closely mirror the phenotype and genotype of primary tumors than do serum-cultured cell lines. Cancer Cell 9: 391-403.
-
Singh SK, Clarke ID, Terasaki M, Bonn VE, Hawkins C, et al. (2003) Identification of a cancer stem cell in human brain tumors. Cancer Res 63: 5821-5828.
-
Pilkington GJ (2005) Cancer stem cells in the mammalian central nervous system. Cell Prolif 38: 423-433.
-
Vescovi AL, Galli R, Reynolds BA (2006) Brain tumour stem cells. Nat Rev Cancer 6: 425-436.
-
Ward RJ, Dirks PB (2007) Cancer stem cells: at the headwaters of tumor development. Annu Rev Pathol 2: 175-189.
-
Pastrana E, Silva-Vargas V, Doetsch F (2011) Eyes wide open: a critical review of sphere-formation as an assay for stem cells. Cell Stem Cell 8: 486-498.
-
Beier D, Hau P, Proescholdt M, Lohmeier A, Wischhusen J, et al. (2007) CD133(+) and CD133(-) glioblastoma-derived cancer stem cells show differential growth characteristics and molecular profiles. Cancer Res 67: 4010-4015.
-
Gunther HS, Schmidt NO, Phillips HS, Kemming D, Kharbanda S, et al. (2008) Glioblastoma-derived stem cell-enriched cultures form distinct subgroups according to molecular and phenotypic criteria. Oncogene 27: 2897-2909.
-
Gavin PR, Fike JR, Hoopes PJ (1995) Central nervous system tumors. Semin Vet Med Surg (Small Anim) 10: 180-189.
-
Thomas WB, Wheeler SJ, Kramer R, Kornegay JN (1996) Magnetic resonance imaging features of primary brain tumors in dogs. Vet Radiol Ultrasound 37: 20-27.
-
Turrel JM, Fike JR, LeCouteur RA, Higgins RJ (1986) Computed tomographic characteristics of primary brain tumors in 50 dogs. J Am Vet Med Assoc 188: 851-856.
-
Louis DN (2006) Molecular pathology of malignant gliomas. Annu Rev Pathol 1: 97-117.
-
Stoica G (2015) Cancer stem cells: current status and future directions. Vet J 205: 124-125.
-
Louis DN, Ohgaki H, Wiestler OD, Cavenee WK, Burger PC, et al. (2007) The 2007 WHO classification of tumours of the central nervous system. Acta Neuropathol 114: 97-109.
-
Brat DJ, Castellano-Sanchez AA, Hunter SB, Pecot M, Cohen C, et al. (2004) Pseudopalisades in glioblastoma are hypoxic, express extracellular matrix proteases and are formed by an actively migrating cell population. Cancer Res 64: 920-927.
-
Lungu GF, Li ML, Xie X, Wang LV, Stoica G (2007) In vivo imaging and characterization of hypoxia-induced neovascularization and tumor invasion. Int J Oncol 30: 45-54.
-
Vaupel P (2004) The role of hypoxia-induced factors in tumor progression. Oncologist 9 Suppl 5: 10-17.
-
Dong S, Nutt CL, Betensky RA, Stemmer-Rachamimov AO, Denko NC, et al. (2005) Histology-based expression profiling yields novel prognostic markers in human glioblastoma. J Neuropathol Exp Neurol 64: 948-955.
-
Rao JS (2003) Molecular mechanisms of glioma invasiveness: the role of proteases. Nat Rev Cancer 3: 489-501.
-
Tysnes BB, Mahesparan R (2001) Biological mechanisms of glioma invasion and potential therapeutic targets. J Neurooncol 53: 129-147.
-
Ossowski L, Aguirre-Ghiso JA (2000) Urokinase receptor and integrin partnership: coordination of signaling for cell adhesion, migration and growth. Curr Opin Cell Biol 12: 613-620.
-
Goldbrunner RH, Bernstein JJ, Tonn JC (1998) ECM-mediated glioma cell invasion. Microsc Res Tech 43: 250-257.
-
Mizejewski GJ (1999) Role of integrins in cancer: survey of expression patterns. Proc Soc Exp Biol Med 222: 124-138.
-
Paulus W, Tonn JC (1994) Basement membrane invasion of glioma cells mediated by integrin receptors. J Neurosurg 80: 515-519.
-
Friedlander M, Brooks PC, Shaffer RW, Kincaid CM, Varner JA, et al. (1995) Definition of two angiogenic pathways by distinct alpha v integrins. Science 270: 1500-1502.
-
Suzuma K, Takagi H, Otani A, Honda Y (1998) Hypoxia and vascular endothelial growth factor stimulate angiogenic integrin expression in bovine retinal microvascular endothelial cells. Invest Ophthalmol Vis Sci 39: 1028-1035.
-
Magnon C, Hall SJ, Lin J, Xue X, Gerber L, et al. (2013) Autonomic nerve development contributes to prostate cancer progression. Science 341: 1236361.
-
Jie Li, Shan Zhu, David Kozono, Kimberly Ng, Diahnn Futalan, et al. (2014) Genome-wide shRNA screen revealed integrated mitogenic signaling between dopamine receptor D2 (DRD2) and epidermal growth factor receptor (EGFR) in glioblastoma. Oncotarget 5: 882-893.
-
Bartek J Jr, Ng K, Bartek J, Fischer W, Carter B, et al. (2012) Key concepts in glioblastoma therapy. J Neurol Neurosurg Psychiatry 83: 753-760.
-
Ng K, Kim R, Kesari S, Carter B, Chen CC (2012) Genomic profiling of glioblastoma: convergence of fundamental biologic tenets and novel insights. J Neurooncol 107: 1-12.
-
Furnari FB, Fenton T, Bachoo RM, Mukasa A, Stommel JM, et al. (2007) Malignant astrocytic glioma: genetics, biology and paths to treatment. Genes Dev 21: 2683-2710.
-
Verhaak RG, Hoadley KA, Purdom E, Wang V, Qi Y, et al. (2010) Cancer Genome Atlas Research Network. Integrated genomic analysis identifies clinically relevant subtypes of glioblastoma characterized by abnormalities in PDGFRA, IDH1, EGFR and NF1. Cancer Cell. 17: 98-110.
-
Minervini MG, Covelli V, Di Pinto N, Papagni S, Russo F (1985) Brain tumor death risk in mental patients. J Neurosurg Sci 29: 97-100.
