

International Journal of Stem cell Research and Therapy
Impact of New Genome Editing Tools on Ips Cell Based Therapies
Jianbo Wu and Radbod Darabi*
Center for Stem Cell and Regenerative Medicine, University of Texas Health Science Center at Houston, USA
*Corresponding author: Radbod Darabi, Center for Stem Cell and Regenerative Medicine/ The Brown Foundation Institute of Molecular Medicine, The University of Texas Health Science Center at Houston, 1825 Pressler Street SRB 630A, Houston, TX 77030, USA, Tel: 713-500-3411; Fax: 713-500-2424; E-mail: radbod.darabi@uth.tmc.edu
Int J Stem Cell Res Ther, IJSCRT-1-002, (Volume 1, Issue 1), Review Article; ISSN: 2469-570X
Received: September 18, 2014 | Accepted: December 11, 2014 | Published: December 14, 2014
Citation: Wu J, Darabi R (2014) Impact of New Genome Editing Tools on iPS Cell Based Therapies. Int J Stem Cell Res Ther 1:002. 10.23937/2469-570X/1410002
Copyright: © 2014 Wu J, et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Abstract
Embryonic stem cells (ES cells) have unique differentiation and self- renewal potential which makes them a potential candidate for cell therapy in degenerative disorders. However, the limitation of deriving ES cells from embryos and their immune-incompatibility hinder their clinical applications. Fortunately, by introduction of somatic cell reprogramming technology into induced pluripotent stem cells (iPS cells), the limitations of ES cells have been circumvented. Therefore iPS cells have been considered as a promising cell source for clinical application in regenerative medicine.
However, in many cases, disease-specific iPS cells need genome modification to replace/ repair the defective gene and regain normal functions before any application. For this reason, Homologous recombination (HR) method is one of the best available options which allow proper gene editing without any undesirable side-effects associated with random integration based methods. However, the efficiency of traditional HR in mammalian cells is extremely low which makes it ineffective for gene correction in human iPS cells.
Fortunately with the recent advancement of new gene editing tools which allow site specific DNA targeting, HR efficiency in iPS cells have been significantly improved. Among these new tools, ZFNs, TALENS and CRISPRs are the most promising ones. This editorial briefly reviews their advantages and shortcomings and debates the impact of these new gene editing technologies on the future of iPS based cell therapies.
Keywords
Genome editing; Human iPS cells; iPS cell therapy; Gene therapy; Induced pluripotent stem cells
Cell Therapy and Stem Cells
Stem cells are undifferentiated cells which can self-renew and differentiate into any specialized cell types. Based on their sources, stem cells from human tissues can be classified as embryonic stem cells (ES cells) and adult stem cells. ES cells isolated from human blastocyst-stage embryos have the capacity for unlimited replication and can be differentiated into all the cell types in human body [1]. However, human adult stem cells derived from non-embryonic tissues are multipotent and can differentiate into only limited number of cell types. Therefore, transplantation of stem cells or their derivatives, known as stem cell therapy, have been considered as one of the most promising approaches to restore the damaged tissues or organs in case of many human diseases such as degenerative disorders [2-4].
However, human ES cells and adult stem cells have limitations which impede their clinical application. Among these, ethical issues and potential immunologic rejection are major obstacles which limit clinical application of human embryonic stem cells [2,4]. On the other hand, in the case of adult stem cells, only limited numbers exist in the adult tissues and they generally have poor expansion/self-renewal potentials [5]. Therefore obtaining large quantity of these cells for transplantation is a big challenge. Furthermore, they also carry immune compatibility issues in case of allogeneic transplants. For example, even in the case of hematopoietic stem cell (HSC) transplantation, the most successful example of cell therapy, over a third of patients who are eligible will not find a suitable human leukocyte antigen (HLA)-matched donor [6]. Fortunately, induced pluripotent stem cell (iPS cell) technology overcomes many of these obstacles and has opened a new era in cell therapy for many genetic and degenerative human disorders.
iPS Technology Evolution
In 2007 the groups of Yamanaka's and Thomson's successfully transformed human fibroblasts into human induced pluripotent stem cells (iPS cells) by over-expression of four transcription factors; Oct4, Sox2, Klf4, and c-Myc or Oct4, Sox2, Nanog, and Lin28 [7,8]. These iPS cells are similar to ES cells in most aspects, including self-renewal and differentiation potential. Therefore, Patient-derived iPS cells offer a promising solution to the major obstacles of stem cell therapy, such as the ethical concerns and immune rejection. This was a great breakthrough for science community as it can potentially provide individualized stem cell therapy for the patients [9].
However, initially both groups had applied retrovirus or lentivirus system for transgene expression, which resulted in the random integration of exogenous genes in the host genome. Even though the expressions of viral transgenes were silenced after complete reprogramming, they still had chances to be re-activated or lead to tumor formation due to random transgene integration into genome [10].
In order to solve these problems, researchers have developed several non-integrative approaches to generate safe iPS cells. Fusaki et al. used Sendai viral vectors to generate iPS cell lines and successfully obtained iPS clones without viral RNA. However, since these viral vectors replicate constitutively, it is still difficult to remove them from host cells and isolate transgene-free clones, even at late passages [11]. Several other research groups have developed episomal vectors (such as minicircle vectors) and have applied transfection to deliver reprogramming factors into target cells [12]. Though in these cases transgene-free iPS clones could be obtained, but still the efficiency is quiet low.
In another attempt to improve the safety of iPS cells, Warren et al. have recently used synthetic modified mRNAs for transgene expression in somatic cells and highly efficient iPS conversion was achieved after 16-days post transfection [13]. Also two other recent studies have successfully used proteins fused with a cell membrane penetrating tag to generate iPS cells [14,15]. Although the mRNA- and protein-based methods are laborious and not easy to handle, human iPS cells generated via these approaches are potentially safer for clinical application. Overall, the recent studies have demonstrated the safe preparation of iPS cells from somatic cells suitable for regenerative purposes.
iPS Cells for Disease Modeling and Regenerative Medicine
So far, the iPS technology has been widely used for disease modeling such as heart disease (e.g. long-QT syndrome), nervous system diseases (e.g. Parkinson's disease), type I diabetes and many other genetic diseases [16-18]. However, its application in regenerative medicine lags far behind due to problems associated with gene modification/correction.
Indeed iPS cell lines derived from patients with genetic diseases still contain disease-associated mutations and display disease-specific phenotypes. Therefore, in case of genetic disorders, gene correction and gain-of-function tests are necessary before application of patient derived iPS cells for clinical applications. Moreover, in some cases, genome modification can also render patient-derived iPS cells with some special function for therapeutic purposes. For example, Monocytes/Macrophages with homozygous deletions of 32-bp sequences (CCR5?32) in C-C chemokine receptor type 5 gene- a major co-receptor used by HIV-1 virus to infect T cells, macrophages, and other cell types- can resist HIV-1 infection [19]. Therefore, gene editing is a necessary tool for altering iPS cells to a favorable cell therapy for many diseases not limited to only genetic disorders.
Gene Editing in iPS Cells
There are two major strategies for the genome modification of patient-derived iPS cells: random integration via viral vectors and site-specific correction via homologous recombination (HR). Although gain-of-function can be achieved through correction of disease-specific iPS cells by viral vectors, genome modifications via viral vectors lead to random integration of viral genes into the host cell genome and have uncertain risks, including tumor formation [20]. This is the major obstacle for using viral vectors for gene modification in iPS cells.
Therefore, the most promising strategy is site-specific genome modification via homologous recombination (HR) mechanism. As HR is a nucleotide sequence exchange between two similar or identical DNA fragments, site-specific genome modification can be achieved by gene targeting vectors with correct DNA replacements between two homology arms. Unfortunately, the efficiency of traditional HR in mammalian cells is extremely low. However, introduction of site specific DNA double-strand breaks (DSBs) can significantly increase the efficiency of HR [21].
Recently, a number of genome editing technologies have been developed to introduce site-specific DSB, including zinc-finger nucleases (ZFNs), transcription activator-like effector nucleases (TALENs) and the RNA-guided CRISPR-Cas nuclease system. Here we overview each method and evaluate their applications for iPS cell gene corrections.
Zinc-Finger Nucleases (ZFNs) and iPS Cells
A ZFN is composed of three protein domains: a nuclear localization signal domain at N-terminal, a nuclease domain derived from the FokI restriction enzyme at C-terminal and a DNA-binding zinc-finger protein (ZFP) domain in the center (Figure 1A). The ZFP domain contains a tandem repeat of Zinc-fingers which recognizes three base pairs of DNA sequence [22]. Three fingers is the minimum number for a ZFN subunit to obtain adequate af?nity. Although including more zinc-fingers can increase binding specificity, but it will also decrease the chances to find a suitable target site. ZFN subunits with 3-6 Zinc-fingers have been successfully used to recognize a DNA sequence of 9-18 bp.
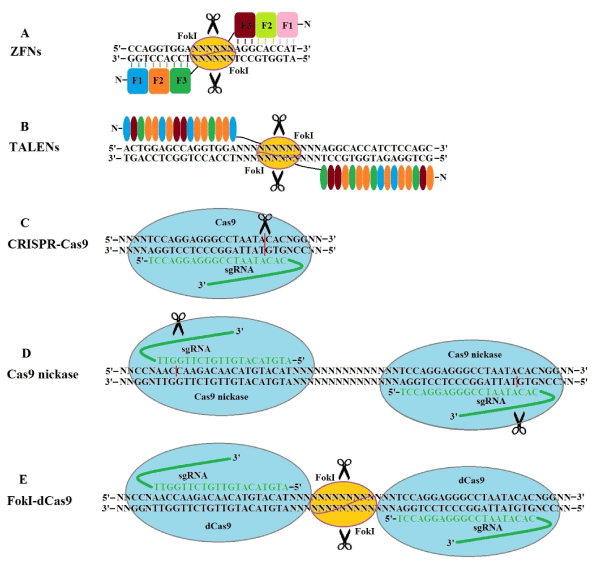 Figure 1: Comparison of different gene editing tools.
Figure 1: Comparison of different gene editing tools.
(A) zinc-finger nucleases (ZFNs). ZFNs recognize two half-sites of the target site with a manner of three base pairs per finger domain.
FokI dimer cleaves a 5-7 bp spacer which links the two half-sites.
(B) Transcription activator-like effector-based nucleases (TALENs). TALENs bind to left and right half-sites in a way of one base pair
per module. The modules with different colors recognize different bases. FokI dimer cleaves a 12-21 bp spacer which separates two
half-sites.
(C) Wild-type Cas9 nuclease. The Cas9 nuclease cleaves the target site of DNA under the direction of a 20 bp short guide RNA
(sgRNA), which recognizes the target site via Watson-Crick base pairing.
(D) Cas9 nickase system. Guided by a pair of sgRNA, Cas9 nickases can efficiently create double-stranded DNA breaks via generation
of double nicking with 5' overhangs (optimally no more than 22 bp).
(E) FokI-dCas9 system. A pair of sgRNAs directs FokI-dCas9 to the left and right half-sites. Dimerization of FokI domains generates
DNA breaks within a 13-25 bp spacer between two half-sites.
View Figure 1
Furthermore, dimerizations of FokI domains are required for nuclease activity of this system. When two ZFN monomers bind DNA in opposite orientations and have an appropriate spacer to permit FokI dimer formation, they form an active endonuclease complex and create DSBs in the genome [23]. DNA DSBs are usually repaired by two major pathways: the non-homologous end joining (NHEJ) or homology-directed repair (HDR) [24].
In the absence of a DNA template, two ends of DSBs are re-ligated through the NHEJ pathway and insertion/deletion (indel) mutations are introduced at the repair sites. In the presence of a repair template, DSBs can be fixed through the HDR pathway [24]. With the combination of ZFNs and gene targeting constructs with a desired sequence flanked by two homology arms, researchers can introduce defined modifications at the interest target sites or correct the mutations in the genome. This ability makes ZFNs as a potential tool for iPS genome editing and repair.
However, there are few limitations for this technology. For instance, although many of individual module fingers which target most of the possible 64 triplet-binding sites have been identified, but it is still difficult to collect 64 zinc- fingers that cover all possible combinations of triplet sites. Therefore, it is still a challenge to generate ZFNs for all the target sequences. Furthermore, even if ZFNs can be successfully generated, some of ZFNs might not cleave genomic DNA efficiently [25]. Moreover, the off-target mutations introduced by ZFNs can lead to toxicity in host cells [26]. Additionally, design and construction of ZFNs is complex and time-consuming [27] as ZFNs are assembled by different zinc- fingers with similar structures. This makes it a complex and time-consuming process to generate ZFNs.
Despite these limitations, ZFNs have been used for gene correction of a number of patient-derived iPS cells. Zou et al. performed ZFNs-assisted gene correction of the β-globin mutation in the iPS cell lines derived from patients with sickle cell anemia [28]. They have successfully conversed one of two mutated β-globin alleles to the wild-type. Unfortunately, after differentiation of gene-corrected heterozygous iPS cells into erythrocytes, the erythroid cells expressed the corrected allele only at the level of 25%-40% of the mutated allele. The remaining loxp sequences or a genetic mutation in HBB 3'-enhancer were speculated to be responsible for the low expression of the corrected allele [28]. ZFNs-mediated gene correction of the iPS cell lines from patients with β-thalassemia and a(1)-antitrypsin deficiency have also been reported recently [29,30]. These studies have confirmed the gene correction by DNA sequencing as well as in vitro and in vivo gain-of-function experiments. More importantly, in the case of a(1)-antitrypsin deficiency, the authors have sequenced exomes of the corrected iPS cells and its parental fibroblasts and did not detect any mutations introduced by off-target events [30]. Overall, despite of their hurdles, ZFNs are still considered as a practical tool for gene modification in iPS cells.
Transcription Activator-Like Effector Nucleases (TALENs) and iPS Cells
Like ZFNs, TALENs (Figure 1B) also contain three functional domains. But different from ZFNs, the DNA-binding domains of TALENs are derived from a class of naturally occurring DNA binding proteins, known as TALEs. TALEs consist of tandem repeats of 33-35 amino acids, which recognize DNA sequence in a way of one repeat to one base pair. These repeats are highly conserved in amino acid sequence, except the amino acids at positions 12 and 13, which are called repeat variable diresidues (RVDs). The residues of RVDs determine the nucleotide binding specificity of each repeat [31].
In the original repeats arrays, four RVDs have been identified: Asn-Asn, Asn-Ile, His-Asp and Asn-Gly, which recognize guanine, adenine, cytosine and thymine, respectively [32]. As mentioned before, the FokI domain functions as a dimer. Therefore, it requires two TALEN monomers with proper orientation and spacing work together to achieve DNA cleavage. The one-to-one correspondence between repeats and base pairs make it possible to target almost any DNA sequence. Although it has been recently reported that the presence of 5-methylated cytosine at the target sites attenuate the binding affinity of TALENs, the replacement of His-Asp RVD repeat with an Asn-Gly RVD repeat can overcome this problem [33].
Consisting of only four kinds of RVD codes, TALENs can be generated within a few days which make them more attractive than ZFNs. Therefore, researches have applied it for the gene correction of disease-specific iPS cells. Ma et al. utilized TALENs to facilitate the gene correction of β-globin mutation in iPS cells from β-thalassemia patients [34]. The authors demonstrated that the gene correction did not cause off-target mutations and erythroblasts derived from corrected iPS cells expressed normal β-globin. TALENs-assisted gene correction was also successfully used to correct the NPC1 mutation in iPS cells derived from patients with Niemann-Pick type C (NPC) disease [35]. The disease-associated defects in NPC1-deficient human hepatic and neural cells were rescued in the cell types derived from corrected iPS cells. Overall, TALENs easier design compared to ZFNs makes them a favorable tool for gene correction in iPS cells.
RNA-Guided CRISPR-Cas Nuclease System and iPS Cells
Most recently, a class of RNA-guided engineered nucleases have been developed from microbial adaptive immune systems named as clustered regularly interspaced short palindromic repeats (CRISPR)/CRISPR-associated (Cas) systems [36,37]. These systems are used by bacteria and archaea to cleave foreign genetic elements. There are three types (I-III) of CRISPR/Cas systems [38]. Due to its simplicity, type II has been well characterized and used for genome engineering [39,40]. This system (Figure 1C) consists of three components: the nuclease Cas9, the CRISPR RNA (crRNA) array and a trans-activating crRNA (tracrRNA) which is required for the maturation of crRNA.
The crRNA array is a long array of direct DNA repeats separated by short variable sequence spacers. These spacers encode guide RNAs, which direct Cas9 to the target DNA through Watson-Crick base-pairing interaction. As the crRNA array can have more than one guide RNAs, targeting multiple sites at once can also be achieved. crRNA and tracrRNA form an active nuclease which this complex recognizes the protospacer-adjacent motif (PAM) site and initiates the crRNA-DNA interaction. In other words, the Cas9 system targets a 20-bp DNA sequence followed by a PAM site at its 3' end. The most popular Cas9, Streptococcus pyogenes cas9 (SpCas9) binds to the 5'-NGG-3' PAM sequence, and 5'-NAG-3' with a lower frequency, while the Cas9 derived from other species recognize different PAM sequences [41].
Cas9 contains two nuclease domains: RuvC and HNH, each of which cuts a different DNA strand and generate double strand breaks (DSBs) together. In order to improve the on-target specificity of Cas9 nuclease, two different kinds of strategies have been developed. In first strategy, a Cas9 nickase that cuts a single strand of DNA was generated by inactivating RuvC nuclease domain via point mutation (D10A) [42,43]. Directing by a pair of guide RNAs, which their target sites have proper space and orientation, the Cas9 nickase can generate a DSB efficiently (Figure 1D). As this strategy a) doubles the length of target sequence and b) the DNA single-strand breaks created by the nickases at off-target sites can be repaired via the high-fidelity base excision repair (BER) pathway, the nicking strategy improves target specificity by up to 1500 time relative to the wild-type Cas9 [42]. It has also been shown that gRNAs with 2-3 bp shorter sequence could further reduce off-target effects induced by Cas9 nickases, which might be a result of more sensitivity to mismatches [44].
In the second strategy, a catalytically inactive Cas9 was also generated and fused with a FokI nuclease domain (Figure 1E). This FokI-dCas9 nuclease was guided by a single gRNA. As The FokI domains must dimerize to acquire nuclease activity, the cleavage activity of FokI-dCas9 requires the binding of two gRNAs to DNA with a defined spacing and orientation [45,46]. Tsai et al. [45] reported that FokI-dCas9 directed by 12 single gRNAs induced indels at the target site with a low frequency of 0.15%, which might be due to recruitment of two FokI-dCas9 molecules to the target locus. Although in this case the frequency of indel mutations induced by a pair of FokI-dCas9 nucleases was indistinguishable from background at five known off-target sites, the chances of mutagenesis induced at other highly match off-target sites remains unknown.
From the practical standpoint, generation of Cas9 nucleases is very simpler than two previous methods as it only needs a pair of oligonucleotides and sub-cloning into the expression vectors of Cas9 systems. This makes Cas9 system as the most popular genome editing tool so far [41]. Therefore many groups are currently using this system for iPS genome editing in iPS cells.
Xie et al. [47] have used CRISPR/Cas9 system to efficiently correct the hemoglobin B (HBB) mutations in iPS cells from β-thalassemia patients. No off-target sites were detected at all six potential off-target sites in the corrected iPS cells. The cells retained their pluripotency and restored expression of HBB after differentiation into erythroblasts. Ye et al. used TALENs- and Cas9-mediated HR to generated genetically homozygous CCR5?32 modified iPS cells with almost 100% efficiency of HR for a single allele and about 22% and 33% for double alleles. There were no detectable off-target mutations in the corrected iPS cells. More importantly, the monocytes/macrophages derived from these modified iPS cells showed resistance to HIV-1 infection [19]. Overall, due to its simple design and target specificity, it seems Cas9 system rapidly takes over the future of genome editing in iPS cell technology.
Concluding Remarks
So far, ZFNs, TALENs and CRISPR-Cas nucleases have been successfully used for genome modification of a number of disease-specific iPS cells, indicating that they are powerful tools for site-specific gene targeting and iPS cell therapy will benefit from them in the future. Among these, CRISPR-Cas nucleases have shown more promises for future genome editing in human iPS cells due to their easy and fast design as well as their relative precision- especially in case of Cas9-nickase version- and efficacy for gene editing.
However, the biggest concern about all above mentioned tools is their safety, as all of these genome editing tools generate DSBs at the target sites as well as off-target sites, which will introduce indel mutations. Although in recent studies, only few mutagenesis at off-target site have been reported but most of these studies only examined off-target mutations at few predicted sites. Therefore, chances of mutagenesis at many other off-target sites still remain unknown. The extent of off-target side effects should be fully evaluated before the clinical application of genome editing technology. Meanwhile, further improvements in binding and cleavage specificity of the genome editing tools will minimize the off-target events and enhance their potential therapeutic application. These make the future of iPS cell therapy very exciting and challenging road map to complete.
References
-
Yu J, Thomson JA (2008) Pluripotent stem cell lines. Genes Dev 22: 1987-1997.
-
Cedar SH, Cooke JA, Luo Z, Patel MJ, Minger SL (2007) From embryos to embryonic stem cells: biopolitics and therapeutic potential. Reprod Biomed Online 14 Spec No 1: 67-73.
-
Lerou PH, Daley GQ (2005) Therapeutic potential of embryonic stem cells. Blood Rev 19: 321-331.
-
Zeng X, Rao MS (2006) The therapeutic potential of embryonic stem cells: A focus on stem cell stability. CurrOpinMolTher 8: 338-344.
-
Yabut O, Bernstein HS (2011) The promise of human embryonic stem cells in aging-associated diseases. Aging (Albany NY) 3: 494-508.
-
Brunstein CG, Wagner JE (2006) Umbilical cord blood transplantation and banking. Annu Rev Med 57: 403-417.
-
Takahashi K, Tanabe K, Ohnuki M, Narita M, Ichisaka T, et al. (2007) Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell 131: 861-872.
-
Yu J, Vodyanik MA, Smuga-Otto K, Antosiewicz-Bourget J, Frane JL, et al. (2007) Induced pluripotent stem cell lines derived from human somatic cells. Science 318: 1917-1920.
-
Inoue H, Nagata N, Kurokawa H, Yamanaka S (2014) iPS cells: a game changer for future medicine. EMBO J 33: 409-417.
-
Miura K, Okada Y, Aoi T, Okada A, Takahashi K, et al. (2009) Variation in the safety of induced pluripotent stem cell lines. Nat Biotechnol 27: 743-745.
-
Fusaki N, Ban H, Nishiyama A, Saeki K, Hasegawa M (2009) Efficient induction of transgene-free human pluripotent stem cells using a vector based on Sendai virus, an RNA virus that does not integrate into the host genome. Proc Jpn Acad Ser B Phys Biol Sci 85: 348-362.
-
Yu J, Hu K, Smuga-Otto K, Tian S, Stewart R, et al. (2009) Human induced pluripotent stem cells free of vector and transgene sequences. Science 324: 797-801.
-
Warren L, Manos PD, Ahfeldt T, Loh YH, Li H, et al. (2010) Highly efficient reprogramming to pluripotency and directed differentiation of human cells with synthetic modified mRNA. Cell Stem Cell 7: 618-630.
-
Zhou H, Wu S, Joo JY, Zhu S, Han DW, et al. (2009) Generation of induced pluripotent stem cells using recombinant proteins. Cell Stem Cell 4: 381-384.
-
Kim D, Kim CH, Moon JI, Chung YG, Chang MY, et al. (2009) Generation of human induced pluripotent stem cells by direct delivery of reprogramming proteins. Cell Stem Cell 4: 472-476.
-
Moretti A, Bellin M, Welling A, Jung CB, Lam JT, et al. (2010) Patient-specific induced pluripotent stem-cell models for long-QT syndrome. N Engl J Med 363: 1397-1409.
-
Soldner F, Hockemeyer D, Beard C, Gao Q, Bell GW, et al. (2009) Parkinson's disease patient-derived induced pluripotent stem cells free of viral reprogramming factors. Cell 136: 964-977.
-
Maehr R, Chen S, Snitow M, Ludwig T, Yagasaki L, et al. (2009) Generation of pluripotent stem cells from patients with type 1 diabetes. Proc Natl Acad Sci U S A 106: 15768-15773.
-
Ye L, Wang J, Beyer AI, Teque F, Cradick TJ, et al. (2014) Seamless modification of wild-type induced pluripotent stem cells to the natural CCR5Δ32 mutation confers resistance to HIV infection. Proc Natl Acad Sci U S A 111: 9591-9596.
-
Madonna R (2012) Human-induced pluripotent stem cells: in quest of clinical applications. Mol Biotechnol 52: 193-203.
-
Sorrell DA, Kolb AF (2005) Targeted modification of mammalian genomes. Biotechnol Adv 23: 431-469.
-
Kim YG1, Cha J, Chandrasegaran S (1996) Hybrid restriction enzymes: zinc finger fusions to Fok I cleavage domain. Proc Natl Acad Sci U S A 93: 1156-1160.
-
Urnov FD, Rebar EJ, Holmes MC, Zhang HS, Gregory PD (2010) Genome editing with engineered zinc finger nucleases. Nat Rev Genet 11: 636-646.
-
Kakarougkas A, Jeggo PA (2014) DNA DSB repair pathway choice: an orchestrated handover mechanism. Br J Radiol 87: 20130685.
-
Wilson KA, McEwen AE, Pruett-Miller SM, Zhang J, Kildebeck EJ, et al. (2013) Expanding the Repertoire of Target Sites for Zinc Finger Nuclease-mediated Genome Modification. Mol Ther Nucleic Acids 2: e88.
-
Szczepek M, Brondani V, Buchel J, Serrano L, Segal DJ, et al. (2007) Structure-based redesign of the dimerization interface reduces the toxicity of zinc-finger nucleases. Nat Biotechnol 25: 786-793.
-
Carroll D, Morton JJ, Beumer KJ, Segal DJ (2006) Design, construction and in vitro testing of zinc finger nucleases. Nat Protoc 1: 1329-1341.
-
Zou J, Mali P, Huang X, Dowey SN, Cheng L (2011) Site-specific gene correction of a point mutation in human iPS cells derived from an adult patient with sickle cell disease. Blood 118: 4599-4608.
-
Wang Y, Zheng CG, Jiang Y, Zhang J, Chen J, et al. (2012) Genetic correction of β-thalassemia patient-specific iPS cells and its use in improving hemoglobin production in irradiated SCID mice. Cell Res 22: 637-648.
-
Yusa K, Rashid ST, Strick-Marchand H, Varela I, Liu PQ, et al. (2011) Targeted gene correction of α1-antitrypsin deficiency in induced pluripotent stem cells. Nature 478: 391-394.
-
Mussolino C, Morbitzer R, Lutge F, Dannemann N, Lahaye T, et al. (2011) A novel TALE nuclease scaffold enables high genome editing activity in combination with low toxicity. Nucleic Acids Res 39: 9283-9293.
-
Deng D, Yan C, Pan X, Mahfouz M, Wang J, et al. (2012) Structural basis for sequence-specific recognition of DNA by TAL effectors. Science 335: 720-723.
-
Deng D, Yin P, Yan C, Pan X, Gong X, et al. (2012) Recognition of methylated DNA by TAL effectors. Cell Res 22: 1502-1504.
-
Ma N, Liao B, Zhang H, Wang L, Shan Y, et al. (2013) Transcription activator-like effector nuclease (TALEN)-mediated gene correction in integration-free β-thalassemia induced pluripotent stem cells. J Biol Chem 288: 34671-34679.
-
Maetzel D, Sarkar S, Wang H, Abi-Mosleh L, Xu P, et al. (2014) Genetic and Chemical Correction of Cholesterol Accumulation and Impaired Autophagy in Hepatic and Neural Cells Derived from Niemann-Pick Type C Patient-Specific iPS Cells. Stem Cell Reports 2: 866-880.
-
Ishino Y, Shinagawa H, Makino K, Amemura M, Nakata A (1987) Nucleotide sequence of the iap gene, responsible for alkaline phosphatase isozyme conversion in Escherichia coli, and identification of the gene product. J Bacteriol 169: 5429-5433.
-
Jansen R, Embden JD, Gaastra W, Schouls LM (2002) Identification of genes that are associated with DNA repeats in prokaryotes. Mol Microbiol 43: 1565-1575.
-
Haft DH, Selengut J, Mongodin EF, Nelson KE (2005) A guild of 45 CRISPR-associated (Cas) protein families and multiple CRISPR/Cas subtypes exist in prokaryotic genomes. PLoS Comput Biol 1: e60.
-
Mali P, Yang L, Esvelt KM, Aach J, Guell M, et al. (2013) RNA-guided human genome engineering via Cas9. Science 339: 823-826.
-
Cong L, Ran FA, Cox D, Lin S, Barretto R, et al. (2013) Multiplex genome engineering using CRISPR/Cas systems. Science 339: 819-823.
-
Hsu PD, Lander ES, Zhang F (2014) Development and applications of CRISPR-Cas9 for genome engineering. Cell 157: 1262-1278.
-
Ran FA, Hsu PD, Lin CY, Gootenberg JS, Konermann S, et al. (2013) Double nicking by RNA-guided CRISPR Cas9 for enhanced genome editing specificity. Cell 154: 1380-1389.
-
Mali P, Aach J, Stranges PB, Esvelt KM, Moosburner M, et al. (2013) CAS9 transcriptional activators for target specificity screening and paired nickases for cooperative genome engineering. Nat Biotechnol 31: 833-838.
-
Fu Y, Sander JD, Reyon D, Cascio VM, Joung JK (2014) Improving CRISPR-Cas nuclease specificity using truncated guide RNAs. Nat Biotechnol 32: 279-284.
-
Tsai SQ, Wyvekens N, Khayter C, Foden JA, Thapar V, et al. (2014) Dimeric CRISPR RNA-guided FokI nucleases for highly specific genome editing. Nat Biotechnol 32: 569-576.
-
Guilinger JP, Thompson DB, Liu DR (2014) Fusion of catalytically inactive Cas9 to FokI nuclease improves the specificity of genome modification. Nat Biotechnol 32: 577-582.
-
Xie F, Ye L, Chang JC, Beyer AI, Wang J, et al. (2014) Seamless gene correction of β-thalassemia mutations in patient-specific iPSCs using CRISPR/Cas9 and piggyBac. Genome Res 24: 1526-1533.
