

International Journal of Stem cell Research and Therapy
Comparative Characterization of Canine and Human Mesenchymal Stem Cells Derived from Bone Marrow
Bertolo A1, Schlaefli P1, Malonzo-Marty C1, Baur M2, Potzel T2, Steffen F3 and Stoyanov J1,4*
1Swiss Paraplegic Research, Nottwil, Switzerland
2Orthopaedics and Spinal Surgery, Swiss Paraplegic Centre, Switzerland
3Section of Small Animal Surgery/Neurology, Vetsuisse Faculty of the University of Zurich, Switzerland
4Institute for Surgical Technology and Biomechanics, University of Bern, Switzerland
*Corresponding author: Jivko Stoyanov, Biomedical Laboratories, Swiss Paraplegic Research, G.A.Zach Strasse 4, CH-6207 Nottwil, Switzerland, Tel: +41 41-939-6635, E-mail: jivko.stoyanov@paraplegie.ch
Int J Stem Cell Res Ther, IJSCRT-2-005, (Volume 2, Issue 1), Research Article; ISSN: 2469-570X
Received: December 10, 2014 | Accepted: January 16, 2015 | Published: January 19, 2015
Citation: Bertolo A, Schlaefli P, Malonzo-Marty C, Baur M, Potzel T, et al., (2015) Comparative Characterization of Canine and Human Mesenchymal Stem Cells Derived from Bone Marrow. Int J Stem Cell Res Ther 2:005. 10.23937/2469-570X/1410005
Copyright: © 2015 Bertolo A, et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Abstract
Bone marrow is a major source of mesenchymal stem cells (MSC), which are used in tissue engineering and other autologous stem cell therapies. Studies designed to use large canine models and translate the results to human practice must take into account the ex vivo and in vitro differences in the bone marrow samples. A set of morphological and physiological markers was used to compare MSC derived from canine and human bone marrow. Despite anti-clotting treatment, frequent bone marrow clotting was a problem with canine samples, so we developed a protocol for enzymatic digestion of undesirable clots. We compared colony forming units (CFU) assay, population doubling time (PDT), senescence-associated β-galactosidase (SA-ß-Gal) activity, as well as the ability of cells to differentiate to osteogenic, adipogenic and chondrogenic phenotypes. Urokinase digestion resulted in recovery of MSC: 4-fold more CFU from canine and 1.6-fold more from human samples when compared to untreated samples. Canine MSC were less robust in vitro - they divided actively only for four weeks in culture, while human cells divided for longer than eight weeks. After six weeks in culture, canine MSC underwent 17 population doublings of while human cells reached 26. The percentage of senescent cells increased linearly with time, but with a faster rate in canine MSC. Human and canine MSC underwent differentiation to all lineages; however canine MSC had generally lower differentiation potential. In conclusion, the discrepancy between canine and human cultures must be considered in future MSC-based therapies based on dogs as animal model.
Keywords
Mesenchymal stem cells, Urokinase, Blood clot, Bone marrow, Stem cell differentiation, Translational medicine
Introduction
Now a days the use of mesenchymal stem cells (MSC) is a corner stone in many translational medicine and tissue engineering strategies. MSC characteristics to differentiate into various cell types [1], migrate, engraft and have anti-inflammatory effects [2,3] reveal their potential for autologous therapies. These cells have found their way into clinical trials for bone and cartilage repair, heart and liver disease as well as for graft versus host disease [4,5].
One of the major challenges in MSC research is to validate the in vitro data by finding good animal models which can mirror the complexity of human disease experience [7]. When animal models work well, for example with mechanistic and evaluation studies of basic biochemistry, physiology and general biology of this approach is feasible. Large dogs are main candidates as potential animal model for human disease research due to their characteristics: (1) relative long life expectancy, (2) large body mass and (3) natural development of diseases that affect human beings. As an example, dogs are also used in studies on heart failure since excitation-contraction coupling processes and the amount of β-myosin heavy-chain isoform are similar to those found in the human myocardium [7]. Or, in the period between 1970 and 2001, dogs were used as animal models in 9% of orthopaedic research studies [8]. In addition, dogs are subjected to natural and environmental factors such as traumatic events, which promote dogs as a suitable animal model to develop MSC-based engineering procedures. For instance, in the field of degenerative intervertebral disc disease, canine models spontaneously develop the disease with ageing and they follow the same medical and surgical treatments as humans [9].
Bone marrow, adipose tissue and the umbilical cord all serve as primary sources for MSC but most promising results have been shown with bone marrow derived stem cells [10], where the common sampling practice is aspiration from the iliac crest [11]. Due to donor variability or withdrawal procedure, MSC yields may vary considerably. It has been shown that the first 5 mL of bone marrow aspirate contain seven to nine times more MSC than the later fractions from withdrawals from a single site of puncture [12]. This drop in bone marrow MSC yield is due to dilution with peripheral blood [12]. Because of the present megakaryocytes and platelets, bone marrow aspirates will clot, unless anticoagulants are mixed with the sample immediately after the aspiration procedure [13,14], but even with anticoagulants, clots may occur.
In our lab we experienced that despite the presence of the anticoagulant sodium citrate, most canine bone marrow samples contained clots and yielded less MSC. To solve this problem, we developed a simple protocol based on an approved thrombolytic drug. Despite the wide use for treatment of patients, only very few publications exist that utilized thrombolytic activities for laboratory applications to rescue clotted samples. In 1987, Niku et al. [15] described the use of streptokinase for dissolving blood clots resulting in functional lymphocytes [15]. More recent publications suggests the use of urokinase [16] and Alteplase [17].
Here, by combining a sequence of tests, our aim was to compare canine and human MSC in vitro and to evaluate their ability to differentiate to chondrogenic, osteogenic and adipogenic lineages. The comparison includes several variables, such as: passage number (which is proportional to in vitro culture age), colony forming unit (CFU) assay, expression of senescence-associated β-galactosidase (SA-β-Gal) marker, cumulative population doublings (CPD) and population doubling time (PDT). We also propose a protocol based on the application of urokinase that may help researchers and medical professionals to make use of bone marrow samples that would else be discarded, thus saving time, material and reducing costs.
Materials and Methods
Collection of human and canine bone marrow aspirates
Human bone marrow aspirates from the iliac crest were collected from consenting donors with the approval of the ethics committee of the canton of Lucerne. Canine bone marrow aspirates from the iliac crest were collected under anesthesia with dog owner's consent.
Isolation of MSC from bone marrow
Human (approx. 20 mL) and canine (approx. 10 mL) bone marrow aspirates were anti-coagulated by addition of 15 mL of 3.8 % sodium citrate (Applichem) immediately after withdrawal (n = 50). After transfer to the laboratory environment, bone marrow extracts were filtered over a 100 μm cell strainer (BD Biosciences).
In case of clot formation, the clot was sliced with a scalpel and triturated with a 1 mL pipet and then treated with urokinase. For the urokinase digest, a 500 μL aliquot of 25 000 Units of urokinase for clinical use (from lyophilized powder 500 000 HS, Medac) was applied directly on the wet clot without any additional buffer, followed by digestion for 1 h at 37°C. After 30 min and after 60 min, the clot was pipetted 10 times using a 1 mL pipet to further disrupt the structure. The sample was again passed over a cell strainer to remove any rests of the clot. Further isolation and plating of MSC depended on the cell source.
Isolation and expansion of canine MSC: After washing the clot rests with PBS, the sample was centrifuged for 10min at 500g. The cell pellet containing erythrocytes and mononuclear cells was resuspended in Alpha-MEM (Gibco), supplemented with 10 % FBS (Bioconcept), 100U/mL penicillin, 100 mg/mL streptomycin, 2.5 mg/mL amphotericin B (all Gibco) and 5ng/mL basic fibroblast growth factor (bFGF; Peprotech). After 3 days culture in a humidified hypoxic (5% oxygen) incubator at 37°C, cells in suspension were removed. Media was changed three times a week until canine MSC were harvested after two weeks of expansion. Along with the setup of the expansion culture, a colony forming assay was set up for quality control. A total of 109 cells (consisting of erythrocytes and mononuclear cells) were plated on 10 cm dishes (Primaria, Falcon). Analogous to the expansion culture, cells in suspension were removed and the dishes were cultured as described. After 14 days, cells were washed with 0.9 % NaCl, fixed with methanol and stained with Giemsa solution (all Applichem).
Isolation and expansion of human MSC: Human MSC were isolated by centrifugation at 800 g for 20 min on a density gradient using H-Lympholyte separation media (Cedarline) in a Leucosep tube (Greiner). After washing, the isolated mononuclear cells were plated in non-hematopoietic stem cell media (Milltenyi). After three days, non-adherent cells were removed and adhering MSC were cultured in DMEM/F12+GlutaMAX, supplemented with 10% FBS, 100 U/mL penicillin, 100 mg/mL streptomycin, 2.5 mg/mL amphotericin B and 5 ng/ml bFGF. For colony formation unit assay, one million mononuclear cells were seeded on 10 cm dishes and stained with Giemsa solution after 14 days in culture.
Senescence Associated Beta-Galactosidase Assay (SA-β-Gal)
After each culture passage (from P2 to P5), both human and canine MSC were seeded at 3'600 cells/cm2 and cultured in the presence of bFGF. After three days, cells were fixed with 2% formaldehyde and 0.2% glutaraldehyde (both Applichem) in PBS and incubated over night at 37°C in a freshly prepared staining solution consisting in 150 mM sodium chloride, 2mM magnesium chloride, 30 mM citric acid/phosphate buffer, 5 mM potassium ferricyanide, 5mM potassium ferrocyanide (all Applichem) and 1 mg/mL 5-bromo-4-chloro-3-indolyl-β-D-galactopyranoside (X-GAL) (Sigma) at pH 6.0 [18]. Following washing with PBS, cells were counterstained with hematoxylin (Invitrogen) and SA-β-Gal positive cells were enumerated by bright field microscopy.
Canine MSC differentiation
Canine MSC from six large dogs (three German shepherd and three Labradors; 6.6 ± 1.4 years old) were differentiated into chondrogenic, osteogenic and adipogenic lineages by stimulation with the appropriate media (cells at Passage 2).
Adipogenic differentiation: MSC were cultured for two weeks in monolayer at 4 × 105 cells/cm2 under two different culture conditions: (1) Adipogenesis maintenance media - DMEM + GlutaMAX, 3% FBS, 100 units/mL penicillin, 100 mg/mL streptomycin, 2.5 μg/mL amphotericin B and 170mM insulin. (all Sigma Aldrich); (2) adipogenesis inducing media - maintenance medium supplemented with 3% FBS, 5% rabbit serum (Gibco), 1 μm dexamethasone, 500 μm 3-Isobutyl-1-methylxanthine (Sigma), 33 μm biotin (Sigma), 5 μm rosiglitazone (Sigma) and 17 μm pantothenate (Sigma) [19]. Lipid droplets were revealed by staining with Oil Red-O (Sigma Aldrich).
Osteogenic differentiation: MSC cultured in monolayer at 7 × 103 cells/cm2 were stimulated for four weeks in Advanced DMEM (GIBCO) + GlutaMAX, 5 % FBS, 100 units/mL penicillin, 100 mg/mL streptomycin, 2.5 μg/mL amphotericin B, 50 μm L-ascorbic acid 2-phosphate, 10 mM β-glycerophosphate (Sigma Aldrich) and 100nM dexamethasone. Calcium mineralization deposits were identified by Von Kossa stain and 5% AgNO3 (Applichem) was reduced to metallic silver.
Chondrogenic differentiation: Cubes (3 mm per side) from a sponge shaped medical device (Biopad, Euroresearch, Milan, Italy), constituted from lyophilized collagen type I, were cut and used as scaffold material to support cellular [20]. MSC were applied to the cubes at 4 × 106 cells/mL (~70 000 cells/cube). Prior to the addition of media, the cells were allowed to adhere to the cubes for 30 min. MSC-collagen constructs were maintained for four weeks in chondrogenic media consisting of DMEM/F12 + GlutaMAX, 2.5 % FBS, 100 units/mL penicillin, 100 mg/mL streptomycin, 2.5 μg/mL amphotericin B, 40 ng/mL dexamethasone (Sigma), 50 μg/mL ascorbic acid 2-phosphate (Sigma), 50μg/mL L-proline (Sigma), 1X Insulin-Transferrin-Selenium (ITS-x, Gibco), and 10 ng/ml transforming growth factor-β1 (TGF-β1, Peprotech). Alcian blue staining was used to visualize accumulation of proteoglycans in constructs' sections.
Human MSC differentiation
Human MSC from seven donors (age: 36 ± 11 years) were differentiated into chondrogenic, osteogenic and adipogenic lineages by stimulation with the appropriate media (cells at Passage 2).
Adipogenic differentiation: MSC were cultured in monolayer at a density of 5 × 105 cells/cm2 for three weeks under two different culture conditions: adipogenesis inducing medium - basal medium (DMEM/F12 + GlutaMAX, 2,5% FBS, 100 units/mL penicillin, 100 mg/mL streptomycin, 2,5 μg/mL amphotericin B) supplemented with 1 μm dexamethasone, 0.5 mM 3-Isobutyl-1-methylxanthine, 0.5 mM indomethacin and 170 mM insulin (all Sigma Aldrich); and adipogenesis maintenance medium - basal medium supplemented with 170 mM insulin. Lipid droplets were revealed by staining with Oil Red-O.
Osteogenic differentiation: MSC cultured for three weeks in monolayer at a density of 5 × 103 cells/cm2 were stimulated for three weeks in DMEM (GIBCO) + GlutaMAX, 5% FBS, 100 units/mL penicillin, 100 mg/mL streptomycin, 2,5 μg/mL amphotericin B, 0.05 mM L-ascorbic acid 2-phosphat, 10 mM β-glycerophosphate and 100 nM dexamethasone. Von Kossa stain was used to identify mineralization deposits in the cell culture.
Chondrogenic differentiation: Collagen cubes were cut and used as scaffold material to support cellular growth. MSC were seeded at a density of 4 × 106 cells/mL in the cubes in 6-well-plates and kept for 30 minutes to allow adhesion, before addition of medium. MSC-collagen constructs were maintained for four weeks in chondrogenic medium consisting of DMEM/F12 + GlutaMAX, 2,5% FBS, 100 units/mL penicillin, 100 mg/mL streptomycin, 2,5 μg/mL amphotericin B, 40 ng/mL dexamethasone, 50 μg/mL ascorbic acid 2-phosphate, 50 μg/mL L-proline, 1X ITS-x, and 10 ng/ml TGF-β1. Alcian blue staining was used to visualize accumulation of proteoglycans in constructs' sections.
Statistical Analysis
Data were expressed as the mean ± SD. Non-parametric Mann-Whitney-Wilcoxon U test for dependent variables was used to determine significant differences between samples. For all tests, p< 0.05 was considered significant. Data analysis was performed with SPSS 14.0 for Windows (SPSS Inc.).
Results
Isolation of MSC from bone marrow aspirates
Despite immediate transfer to anticoagulation solution during isolation of MSC from canine bone marrow, we found that 74% of canine bone marrow samples contained clots, against only 1% of human bone marrow samples (Figure 1). This problem lowered dramatically the yield of MSC isolated due to the loss of all cells trapped by the clot. To increase the cell isolation rate from these samples, we developed a protocol using the thrombolytic drug urokinase.
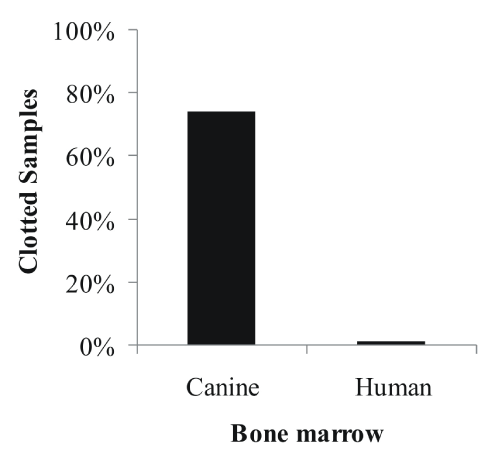
Figure 1: Incidents of clotted canine (n=50) and human (n=50) bone marrow
aspirates after shipment.
View Figure 1
Urokinase digests of bone marrow clots, effects on yield
Since we found that bone marrow clots absorbed a large number of cells, we tested the urokinase protocol on clotted canine and human bone marrow samples to increase MSC yields. After 1 hour incubation in urokinase solution (25 000 U) - applied directly to the clots - the remainder of the clots was weighed (Figure 2A): the clots almost disappeared, with only 15 % (dog) and 9 % (human) of the initial clot weight remaining (p< 0.05).
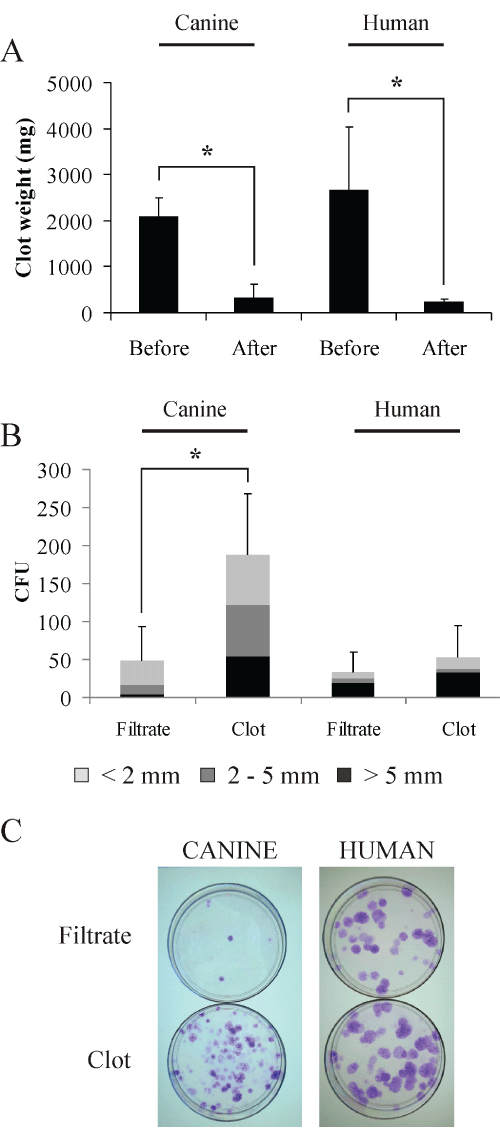
Figure 2: Isolation of human and canine MSC from bone marrow clots
digested with urokinase. Clot weights for canine and human samples before
and after digestion with urokinase (A). The cells recovered from the bone
marrow filtrates and from the digested clots were tested for their ability to form
CFU after two weeks in culture, Giemsa staining (B and C - representative
image). Counted colonies were size class-divided in >5 mm (black), 2-5 mm
(dark grey) and < 2 mm (light grey). (n=5, mean ± SD, * = p ≤ 0.05).
View Figure 2
Further, we analyzed in parallel samples from the bone marrow filtrates and the digested clots by plating in dishes 109 canine cells (erythrocytes included) and 106 human cells. After 2 weeks in culture, colony forming units (CFUs) were visualized by Giemsa staining. We observed a significant 3.8 fold CFU increase (p< 0.05) in urokinase treated canine samples compared to filtrates, i.e. untreated samples where the clot was only removed (Figures 2B,2C). In terms of size, more medium (2-5 mm) and large (>5 mm) colonies were derived from digested canine bone marrow clots (65% of total CFU counts) compared to CFU from filtrates (33% of total counts). Taking into consideration the combined volumes of filtrate and digested clot, the application of the clot-digestion protocol resulted in more than doubling the total yield (2.3 fold) of MSC. Although less prominent, human samples followed a similar trend with 1.6 fold more CFU from treated samples mainly of the large (>5 mm) type, which combining volumes of filtrate and digested clot translated into 1.9 fold increase of the total yield of MSC.
Evaluation of growth kinetics and in vitro senescence between canine and human MSC
In order to characterize MSC senescence, canine and human MSC were cultured in vitro for ~50 days. The number of cumulative population doublings (CPD) showed that canine MSC ceased dividing after 37 days of culture and 17 CPD , while human MSC were still dividing after 47 days and reached 26 CPD (Figure 3A), equivalent to 50% more than canine MSC (p< 0.05).
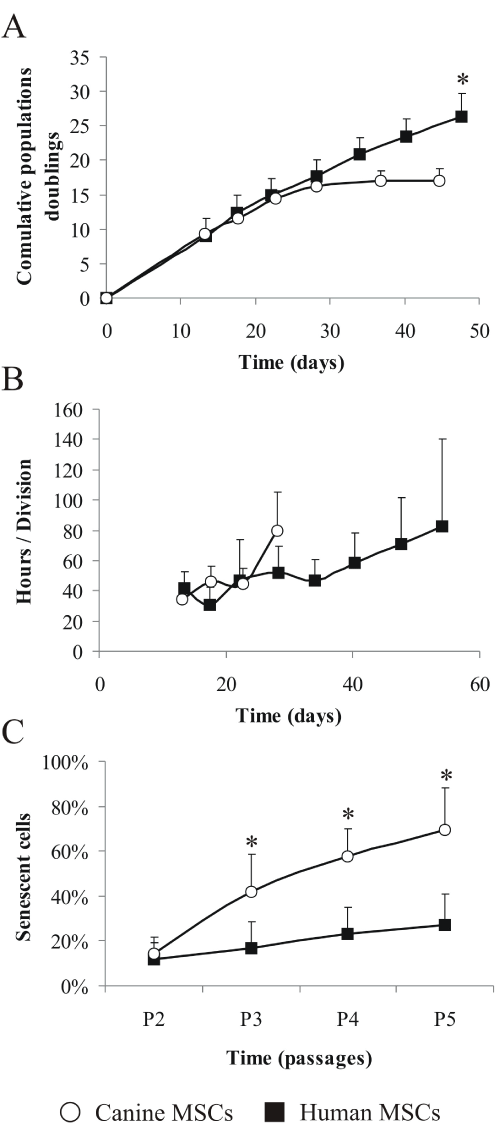
Figure 3: Cell population distinctions between human and canine MSC.
Cumulative population doublings (A) were evaluated at the end of every
passage, as well as population doubling times (B), and calculations were done
based on the time in culture. The number of senescent cells present in MSC
populations (C) from passages P2 to P5 was evaluated by SA-β-gal activity
(canine MSC, n=6, human MSC, n=7; mean ± SD; * = p ≤ 0.05 compared to
canine MSC). White circles=canine MSC; black squares=human MSC.
View Figure 3
We calculated the in vitro population doubling time (PDT): during the initial 28 days of culture human and canine MSC had similar PDT, oscillating between 35 and 45 hours per cell division (Figure 3B) but after 28 days in culture, PDT of canine MSC sharply increased to more than 80 hours per division - a prelude to senescence stage. The same PDT of 80 hours per division human MSC reached only after 54 days in culture.
Finally, MSC were compared for their senescence-associated beta-galactosidase (SA-β-Gal) activity across four in vitro passages, from P2 to P5 (Figure 3C). The number of senescent, SA-β-Gal positive cells, increased with age in vitro and already at P3, the difference in the proportion of senescent cells between canine and human samples was significant (p< 0.05). The average proportion of senescent cells in the canine population of MSC was 14% at P2, but at P3 it increased to 42% and continued to increase for the following passages, reaching 69% at passage 5. Human MSC had less pronounced senescence trend: starting at P2, there were 12% senescent cells which went up only to 27% at P5.
Evaluation of the differentiation potentials of canine and human MSC
To investigate the differentiation potential of canine MSC, we compared them to human MSC (Figure 4). In both species it was possible to differentiate the cells to chondrogenic, adipogenic and osteogenic lineages. Differentiation was histologically assessed with alcian blue, Oil Red O and von Kossa stainings. In canine samples the grade of differentiation observed in the chondrogenic group was lower in comparison to human samples, indicated by lower intensity of the alcian blue staining. The adipogenic lineage of canine MSC produced more numerous but smaller fat vacuoles in comparison with the adipogenic lineage of human origin. Canine MSC which underwent osteogenic differentiation produced few von Kossa positive aggregates, while human MSC deposed mineralized matrix uniformly in the well, overall accumulating larger quantity compared to canine MSC.
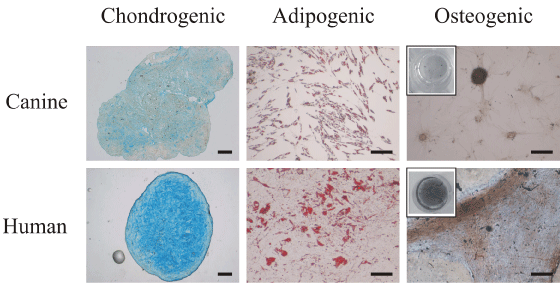
Figure 4: Comparison of differentiation potential of canine (top row) and
human (bottom row) MSC at P2 (Scale bar=200 μm). MSC differentiated to
chondrogenic phenotype were stained by alcian blue, adipogenic phenotype -
stained by Oil Red O, while osteogenesis was revealed by von Kossa staining
(in the inset, pictures of the entire wells).
View Figure 4
Further, we matched the differentiation performance of MSC derived from canine filtrate and clot-digested bone marrow samples, but their ability to differentiate was very similar (data not shown).
Discussion
In this study, we put side by side the in vitro expansion and differentiation potentials of canine and human mesenchymal stem cells (MSC). Such a study was necessary in order to be able to translate results for future therapeutic uses, such as sample handling for tissue engineering approaches in dogs as veterinary patients or as pre-clinical animal model for human trials. Only large dogs (German shepherds and Labradors) were involved in this study, due to their relevant large body mass, ease of obtaining bone marrow material and to avoid inter-breed variations in MSC outputs.
For research and therapeutic purpose, MSC are needed in large numbers and must to be expanded in culture, so the initial number of canine MSC needs to be maximized [21]. When we started to work with canine MSC, we realized that bone marrow aspirates were often clotted upon arrival, while clotted human samples were far less frequent. Since processing of the samples is not done immediately after sampling, resampling to replace clotted specimens would mean a second intervention and consume a lot of resources for patients, healthcare personnel and laboratory staff. Therefore, we tested the idea to replace resampling by the use of urokinase - a clinically approved thrombolytic enzyme - and rescue the cell pool in the clotted samples.
The protocol was tested on clotted human and canine bone marrow samples. One hour incubation was sufficient to digest 85% of the original clot weight in canine samples and over 90% of the human bone marrow clots. The yield of MSC was evaluated by cultivating the filtrate and the clot digest groups for two weeks. Colony forming assays revealed that treatment with urokinase doubled the number of MSC isolated, rescuing cells that otherwise would have been lost. Noteworthy, in our routine work, samples that were digested prior to plating had on average even slightly more CFU than unclotted samples, although a big inter-sample variation was observed due to the heterogeneous donor population [22]. To date, this protocol has been successfully applied on bone marrow clots from different individuals and dogs using urokinase from several lots. Treatment of clotted aspirates for 1 hour at 37°C using 25000 units of clinical grade urokinase improved MSC yield of samples without exception and no apparent detrimental effect in downstream differentiation experiments.
We have isolated canine and human cells from bone marrow aspirates following two distinct protocols, which were previously tested in our lab as the most effective isolation method respectively (data not shown). MSC isolated from dogs also showed dissimilar capacity of growth and differentiation in comparison to human cells. For example the cumulative population doublings of human MSC were significantly higher - by 50% - than canine MSC and while canine and human MSC showed a similar rate of division until day 28, later canine MSC stopped dividing while human cells were still proliferating after day 57. Finally, except at P2 where canine and human MSC had the same proportion of senescent cells, canine MSC populations were significantly more senescent than human and at P5, 70% of canine MSC were senescent against the 25% of senescent cells in the human cell culture. In summary we can conclude that compared to human MSC, canine MSC have significantly shorter life in vitro reflected by the lower number of cell doublings and higher proportion of senescent cells in the population. In addition, dogs had the higher amount of clotted bone marrow samples, almost three out of four against one out of fifty of human samples. The concentration of 5 ng/mL bFGF used to expand cells was chosen based on a dose dependent investigation performed by Wodewotzky et al. On canine MSC demonstrating that high bFGF concentrations (50 ng/mL) had an unexpected inhibitory effect on cell division [23].
MSC can differentiate into cartilage, bone and adipose tissue allowing applications in autologous stem cell therapies. The comparison of the differentiation potentials was based on different differentiation protocols but the state-of-the-art protocols were chosen respectively for canine and human cells. Histology - using von Kossa, Oil Red O and alcian blue stains - showed that human MSC were more productive regarding extracellular matrix accumulation when compared to canine MSC. This contrast was particularly prominent in the differentiation to osteogenic and chondrogenic lineages, while in the adipogenic lineage the divergence were mostly restricted to the size of lipid vacuoles stored by cells. We also investigated the differences in differentiation potential between urokinase treated and unclotted and untreated canine samples, and our analysis showed no significant differences.
Conclusions
Canine MSC can grow and differentiate in vitro, but not as long or as well as human MSC. Our results indicate that the potential of canine MSC to undergo differentiation and to be manipulated in vitro is limited not only in time but also in scale. By using functional biomarkers (number of colony forming units, population doubling time and senescence associated beta-galactosidase assay) to categorize cells derived from bone marrow we showed potentially important differences between canine and human samples and potential consequences for therapeutic use. Furthermore - dealing with sample handling difficulties - we provided a protocol that uses recombinant human urokinase on clotted bone marrow samples to isolate multipotent MSC. This is a directly translatable to human research protocol and a valuable method, since we used an approved pharmaceutical preparation without known antigenicity, so we believe that it can save resources for many research laboratories and has the potential used in therapeutic cell preparations.
From a broader perspective, the correlations between canine and human MSC we described in this study might be helpful for the future development of veterinary practice applications and realistic expectations for the possibility to translate the results to human clinical practice.
Acknowledgments
This work was supported by the Swiss Paraplegic Foundation and Swiss National Foundation Grant CR3I3_140717/1.
References
-
Jiang Y, Jahagirdar BN, Reinhardt RL, Schwartz RE, Keene CD, et al. (2002) Pluripotency of mesenchymal stem cells derived from adult marrow. Nature 418: 41-49.
-
Uccelli A, Moretta L, Pistoia V (2008) Mesenchymal stem cells in health and disease. Nat Rev Immunol 8: 726-736.
-
Bartholomew A, Sturgeon C, Siatskas M, Ferrer K, McIntosh K, et al. (2002) Mesenchymal stem cells suppress lymphocyte proliferation in vitro and prolong skin graft survival in vivo. Exp Hematol 30: 42-48.
-
Wang S, Qu X, Zhao RC (2012) Clinical applications of mesenchymal stem cells. J Hematol Oncol 5: 19.
-
Huey DJ, Hu JC, Athanasiou KA (2012) Unlike bone, cartilage regeneration remains elusive. Science 338: 917-921.
-
Alini M, Eisenstein SM, Ito K, Little C, Kettler AA, et al. (2008) Are animal models useful for studying human disc disorders/degeneration? Eur Spine J 17: 2-19.
-
Lompre AM, Mercadier JJ, Wisnewsky C, Bouveret P, Pantaloni C, et al. (1981) Species- and age-dependent changes in the relative amounts of cardiac myosin isoenzymes in mammals. Dev Biol 84: 286-290.
-
Martini L, Fini M, Giavaresi G, Giardino R (2001) Sheep model in orthopedic research: a literature review. Comp Med 51: 292-299.
-
Bergknut N, Rutges JP, Kranenburg HJ, Smolders LA, Hagman R, et al. (2012) The dog as an animal model for intervertebral disc degeneration? Spine (Phila Pa 1976) 37: 351-358.
-
Baksh D, Song L, Tuan RS (2004) Adult mesenchymal stem cells: characterization, differentiation, and application in cell and gene therapy. J Cell Mol Med 8: 301-316.
-
Malempati S, Joshi S, Lai S, Braner DA, Tegtmeyer K (2009) Videos in clinical medicine. Bone marrow aspiration and biopsy. N Engl J Med 361: e28.
-
Cuthbert R, Boxall SA, Tan HB, Giannoudis PV, McGonagle D, et al. (2012) Single-platform quality control assay to quantify multipotential stromal cells in bone marrow aspirates prior to bulk manufacture or direct therapeutic use. Cytotherapy 14: 431-440.
-
Bain BJ (2001) Bone marrow aspiration. J Clin Pathol 54: 657-663.
-
Goldman JM (1994) A special report: bone marrow transplants using volunteer donors--recommendations and requirements for a standardized practice throughout the world--1994 update. The WMDA Executive Committee. Blood 84: 2833-2839.
-
Niku SD, Hoon DS, Cochran AJ, Morton DL (1987) Isolation of lymphocytes from clotted blood. J Immunol Methods 105: 9-14.
-
Lee YH, Han JY, Seo SY, Kim KH, Lee YA, et al. (2002) Urokinase could increase the yield of progenitor cells during red cell depletion in elapsed and anticoagulated cord blood. Am J Hematol 71: 336-339.
-
St Antoine A, Ketterling MN, Sukov WR, Lowman J, Knudson RA, et al. (2011) Application of thrombolytic drugs on clotted blood and bone marrow specimens to generate usable cells for cytogenetic analyses. Arch Pathol Lab Med 135: 915-919.
-
Dimri GP, Lee X, Basile G, Acosta M, Scott G, et al. (1995) A biomarker that identifies senescent human cells in culture and in aging skin in vivo. Proc Natl Acad Sci U S A 92: 9363-9367.
-
Kisiel AH, McDuffee LA, Masaoud E, Bailey TR, Esparza Gonzalez BP, et al. (2012) Isolation, characterization, and in vitro proliferation of canine mesenchymal stem cells derived from bone marrow, adipose tissue, muscle, and periosteum. Am J Vet Res 73: 1305-1317.
-
Bertolo A, Mehr M, Aebli N, Baur M, Ferguson SJ, et al. (2012) Influence of different commercial scaffolds on the in vitro differentiation of human mesenchymal stem cells to nucleus pulposus-like cells. Eur Spine J 21 Suppl 6: 826-838.
-
Lazarus HM, Haynesworth SE, Gerson SL, Rosenthal NS, Caplan AI (1995) Ex vivo expansion and subsequent infusion of human bone marrow-derived stromal progenitor cells (mesenchymal progenitor cells): implications for therapeutic use. Bone Marrow Transplant 16: 557-564.
-
Bertolo A, Mehr M, Janner-Jametti T, Graumann U, Aebli N, et al. (2013) An in vitro expansion score for tissue-engineering applications with human bone marrow-derived mesenchymal stem cells. J Tissue Eng Regen Med.
-
Wodewotzky TI, Lima-Neto JF, Pereira-Junior OC, Sudano MJ, Lima SA, et al. (2012) In vitro cultivation of canine multipotent mesenchymal stromal cells on collagen membranes treated with hyaluronic acid for cell therapy and tissue regeneration. Braz J Med Biol Res 45: 1157-1162.
