

International Journal of Stem cell Research and Therapy
Enteroendocrine Cells, A Potential Way to Control Intestinal Stem Cell Proliferation
Rihab Loudhaief and Armel Gallet*
Institut Sophia Agrobiotech, France
*Corresponding author: Armel Gallet, Institut Sophia Agrobiotech, UMR INRA 1355/CNRS 7254/UNS, 400 route des Chappes, BP 167, 06903 Sophia Antipolis Cedex, France, Fax: 33 (0)4 92 38 65 87, E-mail: gallet@unice.fr
Int J Stem Cell Res Ther, IJSCRT-3-037, (Volume 3, Issue 2), Perspective; ISSN: 2469-570X
Received: May 27, 2016 | Accepted: June 26, 2016 | Published: July 01, 2016
Citation: Loudhaief R, Gallet A (2016) Enteroendocrine Cells, A Potential Way to Control Intestinal Stem Cell Proliferation. Int J Stem Cell Res Ther 3:037. 10.23937/2469-570X/1410037
Copyright: © 2016 Loudhaief R, et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Perspective
Organisms are subjected during their lifespan to many environmental stresses such as starvation, temperature variations, chemicals, pathogens, injuries... In order to overcome the incoming stress and to yield an adapted answer, organisms have developed throughout evolution robust and conserved mechanisms such as immune response and tissue regeneration helping at maintaining their physiological equilibrium, i.e. their homeostasis. However, the maintenance of the homeostasis can be compromised in certain cases. For example, aging is characterized by an overall decline in tissue homeostasis maintenance with reduced immune function and tissue regeneration capacities and with increased baseline inflammation [1]. Homeostasis can also be disrupted upon chronic or prolonged exposure to toxic or pathogens that can cause damages to organs and tissues [2]. Another example where the homeostasis can be broken down is the predisposition of individuals harboring "silent" mutations to develop pathologies. Under stress conditions, an apparent healthy individual can indeed develop inflammatory diseases, autoimmune syndrome or cancers [3].
The digestive tract is an organ in direct contact with the external milieu facing many xenobiotics or pathogens swallowed along with the food that can harm the mucosa. In the intestine, the replacement of damaged cells relies on intestinal stem cells (ISCs) that divide to give birth to progenitor cells that subsequently differentiate to replenish the gut lining. While under normal condition the intestine is completely renewed in more or less than 2 weeks whether in mouse [4-6] or in Drosophila melanogaster [7,8], upon damages ISC division is accelerated (there is a switch from routine division to proliferation) and the gut is renewed in a couple of days [2]. Many signaling pathways involved in the control of ISC division/proliferation have been identified since the last decade [9,10]). Most of these signals are locally produced (referred to the stem cell niche), coming from neighboring cells such as Paneth cells or surrounding tissues such as the underlying mesenchyme/visceral mesoderm [9-12]. The production of these signals is modulated according to the (local or systemic) need at any given time, allowing the adaptation of ISC division/proliferation to environmental cues [2,13]. This process must be tightly regulated otherwise an uncontrolled ISC proliferation results in overgrowth that can lead to tumor occurrence in the case where ISCs bear a pro-oncogenic mutation.
The enteroendocrine cells (EECs) are secreting cells scattered and interspaced between enterocytes in the gut lining. EECs have long been known to make the link between the nutrient content of the gut and the brain by secreting enteroendocrine peptides that regulate digestive functions and nutritional behavior [14]. More recently, EECs have also been implicated in the local immune response by producing cytokines and anti-microbial peptides [15-18]. EECs are not yet considered as being part of the ISC niche although these cells are indirectly implicated in the control of ISC division (Figure 1A). Indeed it was previously demonstrated that EECs may influence ISC proliferation by secreting peptides regulating the production of growth factors by the niche. In Drosophila it has been shown that the enteroendocrine peptide Tachykinin (ortholog to TAC in mammals) locally secreted by EECs induced the production of Dilp3 (a Drosophila Insulin-like Growth Factors) by the visceral mesoderm [19]. Then Dilp3 promotes ISC proliferation to adapt the growth of the gut to the nutrient availability (Figure 1A) [10]. Still in Drosophila, Scopelliti and colleagues [20] have shown that the enteroendocrine hormone Bursicon produced by EECs is able to constrain ISC division by repressing, in the visceral mesoderm, the expression of the EGF growth factor Vein (Figure 1A). Noteworthy, the Bursicon receptor DLGR2 is the mammalian ortholog of LGR4-6. In mammals, the Glucagon like peptide-2 (GLP-2) secreted by the EECs acts on underlying myofibroblasts to induce the secretion of EGFR/ErbB ligands that further bind receptors in ISCs to stimulate their proliferation (Figure 1A) [21-23]. These data highlight that EECs can act as local regulators of ISC division/proliferation through modulation of the stem cell niche activity in both Drosophila and mice.
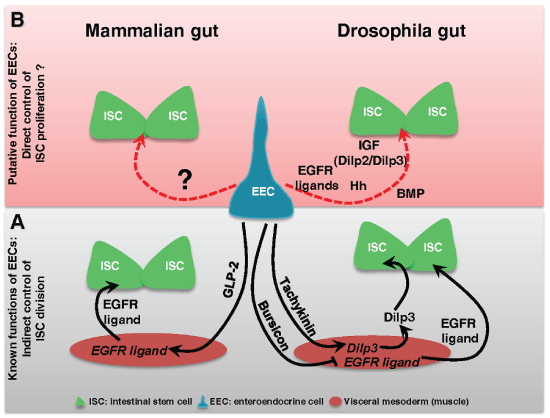
.
Figure 1: Enteroendocrine cells and regulation of intestinal stem cell division.
(A) Scheme summarizing the known functions of EECs in the control of ISC division in mammalian and Drosophila guts. The control is indirect and is mediated by the Glucagon-like peptide-2 (GLP-2) in mammals and by the enteroendocrine hormones Tachykinin and Bursicon in Drosophila; (B) Scheme underlying the putative more direct role of EECs in the control of ISC proliferation upon damages.
View Figure 1
In a recent beautiful transcriptomic analysis in vivo in Drosophila melanogaster, Dutta and Colleagues identified 453 genes that were differentially expressed in EECs upon ingestion of the strong pathogen Pseudomonas entomophila [16]. Among these 453 genes not all are involved in the regulation of the digestive functions, the immune response or paracrine control of the expression of growth factors by the visceral mesoderm. Indeed after our own reading of the data they have released on line (http://flygutseq.buchonlab.com/resources), we found that several genes encoding for secreted factors directly implicated in the control of the ISC proliferation are modulated. In this list of genes, we noticed two Insulin-like Growth Factors (Dilp2 and Dilp3) known to control cell proliferation in many tissues in mammals [24]. In Drosophila, both were already known to control ISC proliferation according to the nutrient availability. Dilp2 is normally released by the central nervous system into the circulation while Dilp3 is locally produced by the visceral mesoderm [10]. We also noticed an increased expression of two EGFR ligands (Keren and Spitz), two IL-6 cytokines orthologs (Upd2 and Upd3), the conserved growth factor Hedgehog (Hh), and two BMP ligands (Dpp and Gbb) (Figure 1B). Whereas EGFR, IL-6/JAK/STAT and Hh signaling pathways positively control ISC division, BMP signaling has a versatile function by either activating or inhibiting ISC division depending of the ligands involved. All these growth factors and cytokines are provided by the niche and they bind to their respective receptors at the surface of ISCs to control their division/proliferation in both vertebrates and Drosophila. Their production is generally increased upon damages to accelerate gut lining regeneration [2,9,10]. Therefore, a potential production of those different factors by EECs [16] can give to these cells an additional important role to maintain gut homeostasis, this time by sending signals directly controlling the behavior of ISCs (Figure 1B). Nevertheless, it will be necessary to investigate whether the production of these factors by EECs is playing a significant role in the control of ISC division before to conclude anything. Because of the high degree of conservation between Drosophila and mammals with respect to the signaling pathways that control ISC division, Drosophila midgut is a suitable model for such an investigation. It is indeed easy to invalidate each candidate in EECs and to assess impacts on ISC division/proliferation at steady state or upon damages. Many investigations in mammals will be also necessary before to definitively unravel the roles of EECs in the control of ISC behavior.
These axes of investigations will probably help at designing cancer therapies specifically targeting EECs. In this perspective, it should be noted that because of the primary role of EECs in the digestive functions, it will be of utmost importance to investigate the relationship between the diet and production of these growth factors by the EECs. Also, identifying which types, where and how EECs control the production of growth factors regulating ISC behavior is essential because of the complexity of the intestine physiology.
References
-
Jasper H (2015) Exploring the physiology and pathology of aging in the intestine of Drosophila melanogaster. Invertebr Reprod Dev 59: 51-58.
-
Bonfini A, Liu X, Buchon N (2016) From pathogens to microbiota: How Drosophila intestinal stem cells react to gut microbes. Dev Comp Immunol 64: 22-38.
-
Sun G, Irvine KD (2011) Regulation of Hippo signaling by Jun kinase signaling during compensatory cell proliferation and regeneration, and in neoplastic tumors. Dev Biol 350: 139-151.
-
Potten CS, Loeffler M (1990) Stem cells: attributes, cycles, spirals, pitfalls and uncertainties. Lessons for and from the crypt. Development 110: 1001-1020.
-
Lopez-Garcia C, Klein AM, Simons BD, Winton DJ (2010) Intestinal stem cell replacement follows a pattern of neutral drift. Science 330: 822-825.
-
Snippert HJ, van der Flier LG, Sato T, van Es JH, van den Born M, et al. (2010) Intestinal crypt homeostasis results from neutral competition between symmetrically dividing Lgr5 stem cells. Cell 143: 134-144.
-
Jiang H, Patel PH, Kohlmaier A, Grenley MO, McEwen DG, et al. (2009) Cytokine/Jak/Stat signaling mediates regeneration and homeostasis in the Drosophila midgut. Cell 137: 1343-1355.
-
Antonello ZA, Reiff T, Ballesta-Illan E, Dominguez M (2015) Robust intestinal homeostasis relies on cellular plasticity in enteroblasts mediated by miR-8-Escargot switch. EMBO J 34: 2025-2041.
-
Vanuytsel T, Senger S, Fasano A, Shea-Donohue T (2013) Major signaling pathways in intestinal stem cells. Biochim Biophys Acta 1830: 2410-2426.
-
Pasco MY, Loudhaief R, Gallet A (2015) The cellular homeostasis of the gut: what the Drosophila model points out. Histol Histopathol 30: 277-292.
-
Biswas S, Davis H, Irshad S, Sandberg T, Worthley D, et al. (2015) Microenvironmental control of stem cell fate in intestinal homeostasis and disease. J Pathol 237: 135-145.
-
Sailaja BS, He XC, Li L (2016) Regulatory niche in intestinal stem cells. J Physiol.
-
Peterson LW, Artis D (2014) Intestinal epithelial cells: regulators of barrier function and immune homeostasis. Nat Rev Immunol 14: 141-153.
-
Latorre R, Sternini C, De Giorgio R, Greenwood-Van Meerveld B (2016) Enteroendocrine cells: a review of their role in brain-gut communication. Neurogastroenterol Motil 28: 620-630.
-
Psichas A, Reimann F, Gribble FM (2015) Gut chemosensing mechanisms. J Clin Invest 125: 908-917.
-
Dutta D, Dobson AJ, Houtz PL, Glaßer C, Revah J, et al. (2015) Regional Cell-Specific Transcriptome Mapping Reveals Regulatory Complexity in the Adult Drosophila Midgut. Cell Rep 12: 346-358.
-
Park JH, Chen J, Jang S, Ahn TJ, Kang K, et al. (2016) A subset of enteroendocrine cells is activated by amino acids in the Drosophila midgut. FEBS Lett 590: 493-500.
-
Katherine Beebe, Dongkook Park, Paul H. Taghert, Craig A. Micchelli (2015) The Drosophila Prosecretory Transcription Factor dimmed Is Dynamically Regulated in Adult Enteroendocrine Cells and Protects Against Gram-Negative Infection. G3 (Bethesda) 5: 1517-1524.
-
Amcheslavsky A, Song W, Li Q, Nie Y, Bragatto I, et al. (2014) Enteroendocrine cells support intestinal stem-cell-mediated homeostasis in Drosophila. Cell Rep 9: 32-39.
-
Scopelliti A, Cordero JB, Diao F, Strathdee K, White BH, et al. (2014) Local control of intestinal stem cell homeostasis by enteroendocrine cells in the adult Drosophila midgut. Curr Biol 24: 1199-1211.
-
Rowland KJ, Brubaker PL (2011) The "cryptic" mechanism of action of glucagon-like peptide-2. Am J Physiol Gastrointest Liver Physiol 301: G1-8.
-
Furness JB, Rivera LR, Cho HJ, Bravo DM, Callaghan B (2013) The gut as a sensory organ. Nat Rev Gastroenterol Hepatol 10: 729-740.
-
Wong VW, Stange DE, Page ME, Buczacki S, Wabik A, et al. (2012) Lrig1 controls intestinal stem-cell homeostasis by negative regulation of ErbB signalling. Nat Cell Biol 14: 401-408.
-
Weroha SJ, Haluska P (2012) The insulin-like growth factor system in cancer. Endocrinol Metab Clin North Am 41: 335-350,
