

Journal of Otolaryngology and Rhinology
In Vitro Modulation of MMP-2 and MMP-9 Secretion by Cytokines, Inducers, and Inhibitors in Head and Neck Squamous Carcinoma Cells (FaDu) and Tongue Carcinoma Cells (SCC-25)
M Waheed Roomi, Bilwa Bhanap, Aleksandra Niedzwiecki* and Matthias Rath
Department of Oncology, Dr. Rath Research Institute, USA
*Corresponding author:
Dr. Aleksandra Niedzwiecki, Department of Oncology, Dr. Rath Research Institute, 1260 Memorex Drive, Santa Clara, CA 95050, USA, E-mail: author@drrath.com
J Otolaryngol Rhinol, JOR-3-029, (Volume 3, Issue 1), Research Article
Received: November 02, 2016 | Accepted: February 18, 2017 | Published: February 21, 2017
Citation: M Waheed Roomi, Bhanap B, Niedzwiecki A, Rath M (2017) In Vitro Modulation of MMP-2 and MMP-9 Secretion by Cytokines, Inducers, and Inhibitors in Head and Neck Squamous Carcinoma Cells (FaDu) and Tongue Carcinoma Cells (SCC-25). J Otolaryngol Rhinol 3:029.
Copyright: © 2017 MW Roomi, et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Abstract
Head and neck squamous cell carcinoma (HNSCC) is a devastating disease with poor prognosis. Matrix metalloproteinases (MMPs), specifically, MMP-2 and MML-9 play a critical role in tumorigenesis by their ability to modify tumor microenvironments, facilitating cell invasion and metastasis. We compared MMP secretion by various cytokines, inducers, mitogen, inhibitors that regulate MMP-2 and MMP-9 activities in FaDu and Tongue cancer cell lines. The cells were grown in appropriate media, at near confluence, they were washed with PBS and inoculated with different concentrations of cytokines (TNF-α, IL-β), mitogen (lipopolysaccharides), inducers (PMA), chemical inhibitors (doxycyline, dexamethosone, actinomycin-D, and cyclohexamide) and natural inhibitors (epigallocatechin gallate, and the nutrient mixture. After 24 hours, the media were removed and analyzed for MMP-2 and MMP-9 by gelatinase zymography and quantitatively analyzed by densitometry. Our results showed that these agents have an up and down regulating effect on MMP-2 and MMP-9 secretion, suggesting the clinical usage of these agents may be beneficial in management of HNSCC cancers.
Keywords
Nutrition mixture, Matrix metalloproteinase's, Tongue carcinoma, HNSCC, Cytokines, Inducers, Inhibitors
Abbreviations
NM: Nutrient Mixture; HNSCC: Head and Neck Squamous Cell Carcinoma; TC: Tongue Carcinoma; MMP: Matrix Metalloproteinases; LPS: Lipopolysaccharides
Introduction
Head and neck squamous cell carcinoma (HNSCC) is a devastating disease with poor prognosis. In the United States, HNSCC is the sixth most commonly diagnosed malignancy and approximately 60,000 cases are diagnosed annually. These include cancers of the oral cavity, larynx, salivary glands, tongue, thyroid, and nose [1,2]. Men are most commonly affected over the age of 50. Head and neck cancers are known for their aggressive nature and propensity to metastasize. The greatest risk factors for HNSCC are tobacco and alcohol use. Eighty-five percent of head and neck cancer cases can be linked to tobacco use including smokeless tobacco [2,3]. Other risk factors for HNSCC include sun exposure, occupational exposure to nickel, radium, chromium, mustard gas, leather, wood dust, and radiation, Epstein Barr Virus infections, and DNA repair disorders such as Fanconi anemia. The resulting costs associated with the treatment of HNSCC in the United States alone add up to billions of dollars per year for surgery, radiation therapy, chemotherapy and post-treatment care [4].
Although the prognosis for successful treatment of HNSCC is good when detected early, over 50% of those with HNSCC seek treatment for advanced stages of the disease when a favorable prognosis decreases significantly [5]. Moreover, radiotherapy and chemotherapy, which are the preferred modes of treatment for HNSCC take a serious physiological toll on the patient [6,7]. Chemotherapy and radiotherapy indiscriminately attack normal as well as diseased cells. Such treatments often lead to serious side effects like anemia, an impaired immune system predisposing the patient to infections, and bleeding and additional cancers caused by the highly toxic nature of chemotherapeutic agents or radiation damage to DNA from radiotherapy [8]. For HNSCC, even when detected and treated in its early stages, the radiation or chemotherapy used creates a 9%-23% incidence of secondary tumors, which decreased overall survival rates [7,8].
Degradation of the extracellular matrix (ECM) by matrix metalloproteinases (MMPs) play a critical role in the formation of tumors and metastasis. MMPs are proteolytic enzymes that digest collagen IV and other components of ECM and the cell basement membrane. In particular MMP-2 and MMP-9 play a key role in the degradation and a good correlation has been observed in increased incidence of invasion and metastasis with increased MMP-2 and MMP-9 [9-11]. Moreover, an increased MMP-9 secretion was noted to correlate with aggressiveness, relapse and shortened survival rates in head and neck cancers [12-14]. The essential role played by the MMPs in metastasis makes MMP-2 and MMP-9 inhibitors (MMPIs), logical targets for therapeutic intervention in cancers. The identification and synthesis of specific non-toxic MMP-2 and MMP-9 inhibitors has been the target of extensive research for several years [15-18].
However, to date the MMPIs that have been clinically tested have not been proven effective in Phase 3 clinical trials because of poor protocols. MMP activity is regulated by, and dependent upon, environmental influences from surrounding stroma cells, ECM proteins, cytokines, inducers and inhibitors, and other factors [19,20]. Furthermore, MMPs are regulated at multiple levels including transcription, modulation of messenger RNA half-life (translation), secretion, localization, activation, and inhibition [21].
In this study, we investigated the effects of selected cytokines, inducers, mitogens and natural and chemical inhibitors, affecting cancer cell metabolism on the regulation of MMP-2 and MMP-9 activities on HNSCC and SC-255 cell lines. We also studied the agents that affect transcription and translation of genes, cyclohexamide and actinomycin-D respectively. Here we also report the dose dependent MMP-2 and MMP-9 inhibitory effects of a non-toxic nutrient mixture (NM) consisting of vitamin C, L-lysine, L-proline, and green tea extract (EGCG) compared to other MMP-2 and MMP-9 inhibitors, cytokines, mitogens, and inducers.
Materials and Methods
Materials
The HNSCC (FaDu), tongue cell carcinoma (SCC-25) cell lines used were purchased from ATCC (American Type Culture Collection, Rockville, MD, USA), along with Dulbecco's Modified Eagle media. Antibiotics such as penicillin, streptomycin, and fetal bovine serum (FBS) were obtained from Gibco (Long Island, NY). Twenty-four well tissue culture plates were obtained from Costar (Cambridge, MA). Gelatinase zymography was performed in 10% Novex precase SDS polyacrylamide gel (Invitrogen Inc.) with 0.1% gelatin in non-reducing conditions. Interleukin 1β (IL-β), tumor necrosis factor-alpha (TNF-α), phorbol 12 myristate (PMA). Lipopolysaccharides (LPS), doxycycline, EGCG, actinomycin-D, retinoic acid, and dexamethasone, were purchased from Sigma (St. Louis, MO). The nutrient mixture (NM), prepared by Vita-Tech was composed of the following ingredients in the relative amounts indicated: Vitamin C (as ascorbic acid and as Mg, Ca, and palmitate ascorbate) 700 mg; L-lysine 1000 mg, proline 750 mg; L-arginine 500 mg; N-acetyl cysteine 200 mg; standardized green tea extract (80% polyphenol) 1000 mg; selenium 30 μg; copper 2 mg; and manganese 1 mg. All other reagents used were of high quality and were obtained from Sigma, unless otherwise noted.
Cell cultures
HSNCC cell lines were grown in Dulbecco's Modified Eagle Media supplemented with 10% FBS, 100 U/ml penicillin, and 100 μg/ml streptomycin in 24 well tissue culture plates. The cells were plated at a density of 1 × 105 cells/ml and grown to confluency in a humidified atmosphere at 5% CO2 at 37 °C. Serum-supplemented medium was removed and the cell monolayer was washed once with phosphate buffered saline (PBS) and once with the recommend serum-free medium. The cells were incubated in 0.5 ml of serum-free medium with various cytokines, inducers, mitogens, and inhibitors in triplicate. Inducer- PMA (10, 25, 50, 100 ng/mL); cytokines- TNF-α (0.1, 1, 10, 25 μg/mL); IL-1β (0.1, 1, 10, 25 μg/mL); mitogen- LPS (10, 25, 50,100 μg/mL); and inhibitors- EGCG (10, 25, 50, 100 μM) with and without PMA; doxycycline (10, 25, 50, 100 μM), with and without PMA; NM (10, 50, 100, 500, 1000 μg/mL) with PMA 100 ng/ml; retinoic Acid (50 μM); dexamethasone (50 μM); and actinomycin-D (2 and 4 μg/mL). The plates were returned to the incubator. After 24 hours, the resulting condition media from each treatment were collected separately, pooled and centrifuged at 4 °C for 10 minutes at 3000 rpm in a microfuge to remove cells and cell debris. The clear supernatant was collected and used for gelatinase zymography as described below.
Gelatinase zymography
Gelatinase zymography was performed in a 10% NOVEX Pre-Cast SDS Polyacrylamide Gel (Invitrogen Corporation) in the presence of a 0.1% gelatin under non-reducing conditions. Culture media (20 μL) were mixed with a sample buffer and loaded for SDS-PAGE with tris-glycine SDS buffer as suggested by the manufacturer (Novex). Samples were not boiled before electrophoresis. Following electrophoresis, the gels were washed twice in a 2.5% Triton X-100 for 30 minutes at room temperature to remove SDS. The gels were then incubated at 37 °C overnight in a substrate buffer containing 50 mM Tris-HCl and 10 mM CaCl2 at pH 8.0 and stained with a 0.5% Coomassie Blue R250 in a 50% methanol and a 10% glacial acetic acid for 30 minutes and destained. Protein standards were run concurrently and approximate molecular weights were determined by plotting the relative mobilities of known proteins. Gelatinase zymograms were scanned using a Canoscan 9950F Canon scanner at 300 dpi. The intensity of the bands was evaluated using the pixel-based densitometer program Un- Scan-It, Version 5.1, 32-bit, by Silk Scientific Corporation (P.O. Box 533, Orem, UT 84059, USA), at a resolution of 1 Scanner Unit (1/100 of an inch for an image that was scanned at 100 dpi).
Results
Effect of inducers, cytokines and mitogens
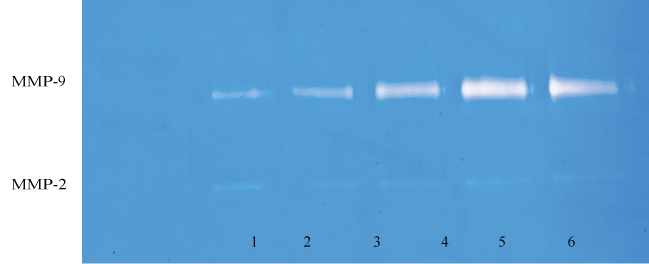
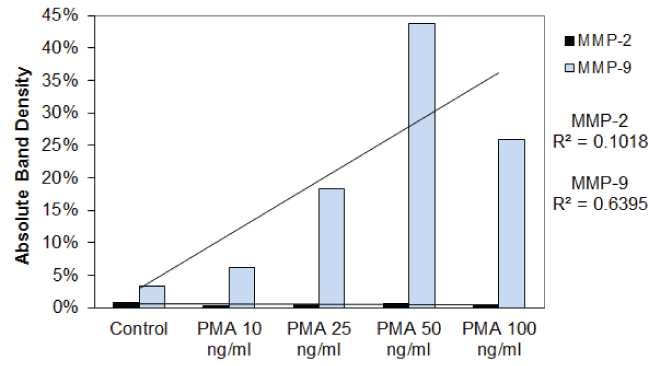
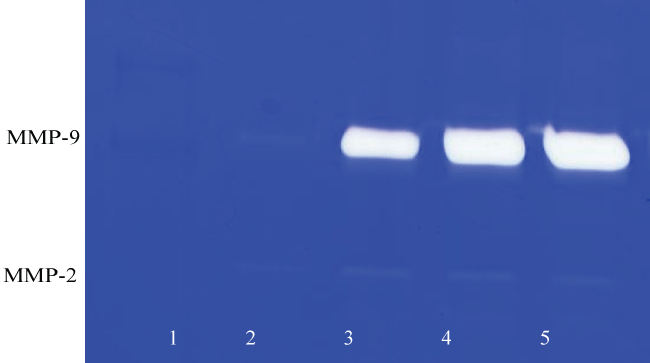
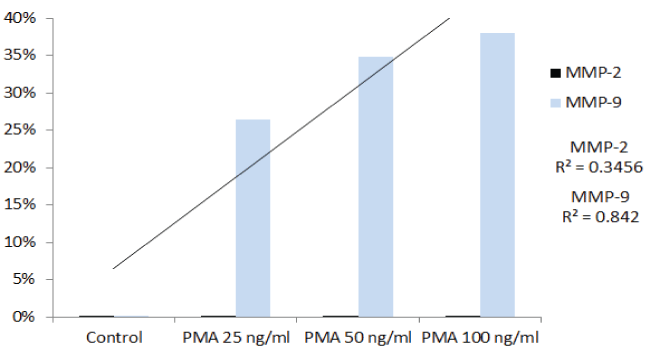
Effect of inducers (PMA), cytokines( TNF-α, IL-1β), and mitogen ( LPS) on MMP-2 and MMP-9 in HNSCC cell line (FaDu) and TC cell line (SCC-25): Table 1 shows the quantitative densitometry results from the effect of inducers PMA, TNF-α, IL-1β and LPS on MMP-2 and MMP-9 secretion in FaDu and TC cell lines. Figure 1A and figure 1C show gelatinase zymography of MMP-2 and MMP-9 secretion in PMA (0-100 ng/ml) treated TC and FaDu cells, respectively, and figure 1B and figure 1D shows the densitometry analysis of their zymograms. As shown in figure 1A, FaDu cells expressed two faint bands, and TC cells expressed two moderate bands corresponding to MMP-2 (72 kDa) and MMP-9 (92 kDa) (figure not shown). On PMA stimulation TC still showed two bands corresponding to MMP-2 and MMP-9. PMA did not alter the pattern of MMP-2 secretion; however, PMA treatment showed a significant dose dependent increase in MMP-9 secretion. PMA had no stimulatory effect on MMP-2 in FaDu cells, and significant dose dependent stimulatory effect on MMP-9 secretion. These results were further confirmed by densitometry analysis (Trend analysis: MMP-2, R2 = 0.3456; MMP-9, R2 = 0.842 for FaDu and MMP-2, R2 = 0.4306; MMP-9, R2 = 0.5337 for TC cell line). TNF-α, IL-1β and LPS have a stimulatory effect on MMP-9 secretion in both FaDu and TC cells in dose dependent manner. TNF-α and IL-1β had a negligible effect on MMP-2 in either cell line, however LPS had a moderate effect on MMP-2 secretion in FaDu and no effect on TC cells.
.
Figure 1: Effect of PMA on MMP-2 and MMP-9 secretion in tongue carcinoma cells (SCC-25) and laryngeal squamous carcinoma (FaDu).
Figure 1A: Gelatinase zymogram of SCC-25 cells treated with PMA shows a dose-dependent increase in MMP-9. Legend: 1-Markers, 2-Control, 3-6: 10, 25, 50, 100 ng/mL PMA.
View Figure 1A
.
Figure 1B: Densitometry analysis of the gelatinase zymogram showing band density as a percentage of the sum of all band densities in the zymogram.
View Figure 1B
.
Figure 1C: Gelatinase zymogram of FaDu cells treated with PMA shows a dose-dependent increase in MMP-9. Legend: 1-Markers, 2-Control/no treatment, 3-5: 25, 50, 100 ng/ml PMA.
View Figure 1C
.
Figure 1D: Densitometric analysis of the gelatinase zymogram showing band density as a percentage of the sum of all band densities in the zymogram.
View Figure 1D
![]()
Table 1: Effect of inducers, cytokines and mitogens on MMP-2 and MMP-9 secretion in laryngeal squamous cells carcinoma cells (FaDu) and tongue cancer cells (SCC-25).
View Table 1
Effect of inhibitors doxycycline and EGCG on MMP-2 and MMP-9 secretion in FaDu and TC cell lines: The effects of doxycycline and EGCG on MMP-2 and MMP-9 secretion in FaDu and TC cells were investigated with and without PMA. As shown in table 2 quantitative densitometry analysis of gelatinase zymography shows a dose dependent decrease in MMP-2 and MMP-9 secretion with doxycycline and EGCG. When stimulated with PMA to express MMP-9, both MMP-2 and MMP-9 decreased in a dose dependent manner with EGCG in the FaDu cell line only (TC not done). Zymography and densitometry graphs of MMP-2 and MMP-9 secretion in doxycycline treated with PMA are shown in figure 2A through figure 2D. Effect of inhibitors, retinoic acid, dexamethasone, actinomycin-D and cyclohexamide on MMP-2 and MMP-9 secretion on FaDu and TC cell lines: As shown in table 3 the secretion of MMP-2 and MMP-9 was significantly inhibited by dexamethasone (50 μM), actinomycin-D (2 and 4 μg/ml), and cyclohexamide (2 and 4 μg/ml), and completely blocked by retinoic acid (50 μM) in the TC cell line. In the FaDu cell line, the secretion of MMP-2 was totally blocked by dexamethasone, actinomycin-D, retinoic acid, and cyclohexamide in a dose dependent manner. MMP-9 secretion in the FaDu cell line was significantly inhibited by dexamethasone, retinoic acid, actinomycin-D and cyclohexamide.

.
Figure 2: Effect of EGCG on MMP-2 and MMP-9 secretion in SCC-25 and FaDu cells.
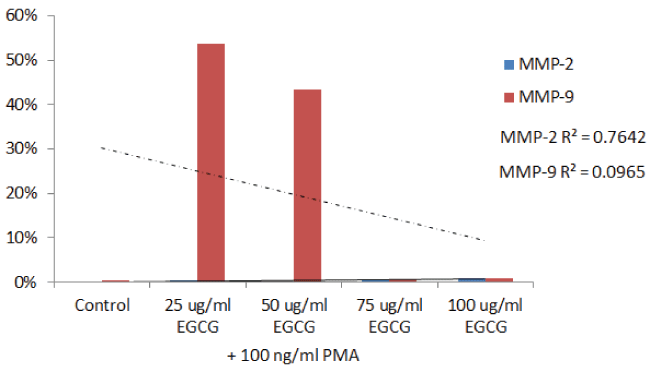
Figure 2A: Gelatinase zymogram of laryngeal squamous cell carcinoma cells treated with EGCG shows a dose-dependent decrease in MMP-2. Legend: 1--Control/no treatment, 2-5: 25, 50, 75, 100 μM EGCG with 100 ng/ml PMA.
View Figure 2A
.
Figure 2B: Densitometric analysis of the gelatinase zymogram of laryngeal squamous cell carcinoma showing band density as a percentage of the sum of all band densities in the zymogram.
View Figure 2B
.
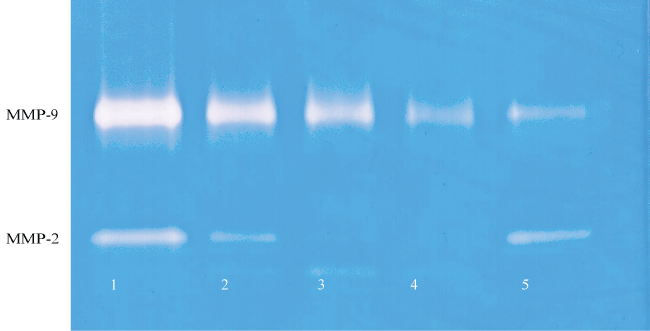
Figure 2C: Gelatinase zymogram of tongue carcinoma cells treated with EGCG shows a dose-dependent decrease in both MMP-2 and MMP-9. Legend: 1-Control/no treatment, 2-5: 25, 50, 75, 100 μM EGCG with 100 ng/ml PMA.
View Figure 2C
.
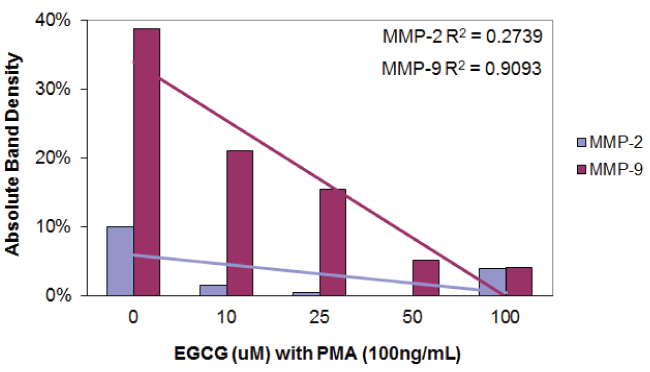
Figure 2D: Densitometric analysis of the gelatinase zymogram of tongue cancer cells showing band density as a percentage of the sum of all band densities in the zymogram.
View Figure 2D
![]()
Table 2: Effect of inhibitors doxycycline and ECGC with and without PMA treatment on laryngeal squamous cell carcinoma (FaDu) and tongue cancer cells (SCC-25).
View Table 2
![]()
Table 3: Effect of inhibitors dexamethasone, actinomycin-D, cyclohexamide and retinoic acid on MMP-2 and MMP-9 secretion in laryngeal squamous cell carcinoma (FaDu) and tongue cancer cells (SCC-25).
View Table 3
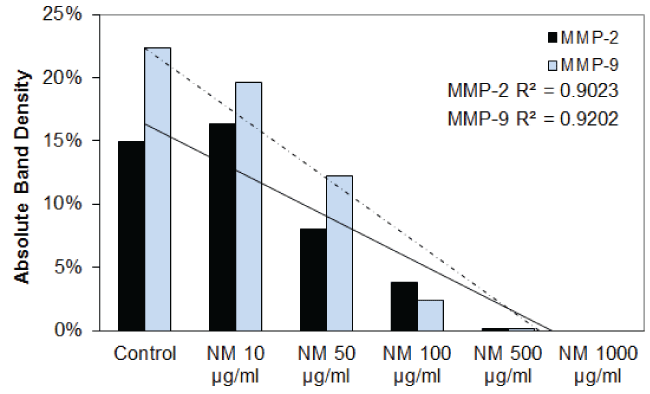
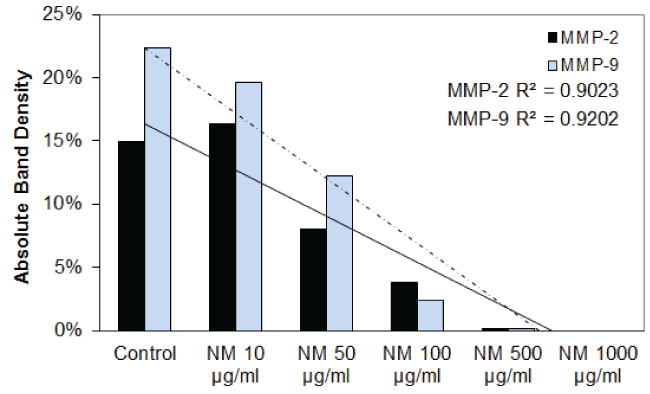
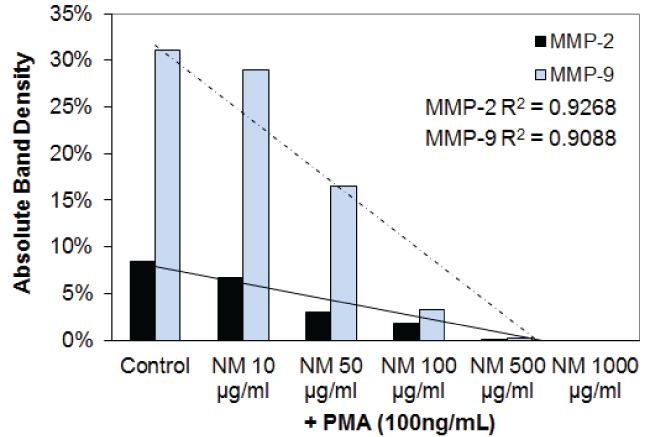
Effect of nutrient mixture (NM) on MMP-2 and MMP-9 secretion in FaDu and TC cell lines: The effect of the nutrient mixture on the secretion of MMP-2 and MMP-9 was also investigated. The results are shown in table 4 and figure 3A through figure 3H. NM showed a dose dependent decrease in MMP-2 and MMP-9 secretion in FaDu, and TC cells. In the PMA stimulated FaDu and TC cells, both MMP-2 and MMP-9 were decreased in a dose-dependent fashion by the NM. The PMA stimulated FaDu and TC cells both showed dose dependent decrease in MMP-2 and MMP-9 secretion caused by NM.

.
Figure 3: Effect of nutrient mixture alone (A-D) and nutrient mixture and PMA (E-H) in squamous cell carcinoma cells (FaDu) and tongue carcinoma cells (SCC-25) on secretion of MMP-2 and MMP-9.
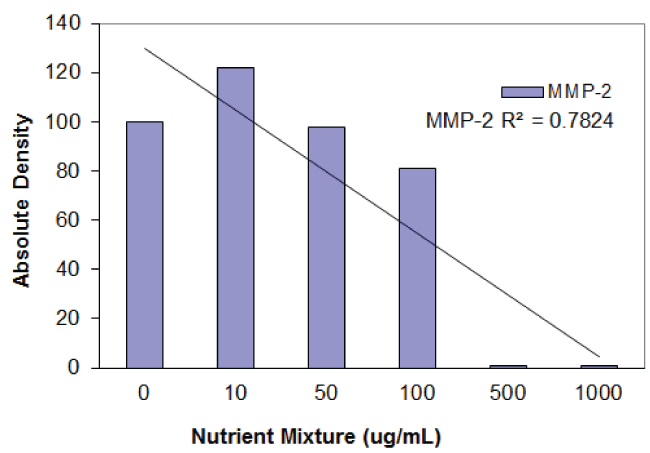
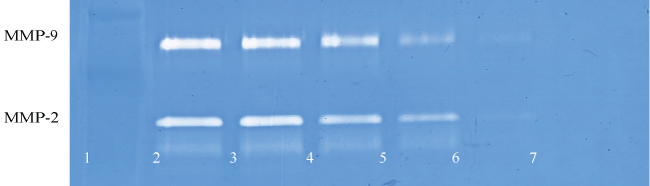
Figure 3A: Gelatinase zymogram of laryngeal squamous cell carcinoma cells treated with a nutrient mixture. Legend: 1- Marker, 2- Control/no treatment, 3-7: 10, 50, 100, 500, 1000 μg/ml Nutrient Mixture.
View Figure 3A
.
Figure 3B: Results of densitometric analysis of the FaDu gelatinase zymogram showing a dose- dependent decrease of MMP-2 secretion after treatment with nutrient mixture.
View Figure 3B
.
Figure 3C: Gelatinase zymogram of tongue carcinoma cells treated with a nutrient mixture. Legend: 1-
Marker, 2-Control/no treatment, 3-7: 10, 50, 100, 500, 1000 μg/ml Nutrient Mixture.
View Figure 3C
.
Figure 3D: Results of densitometric analysis of the SCC-25 gelatinase zymogram showing a dose-dependent decrease in the secretion of both MMP-2 and MMP-9 after treatment with nutrient mixture.
View Figure 3D
.
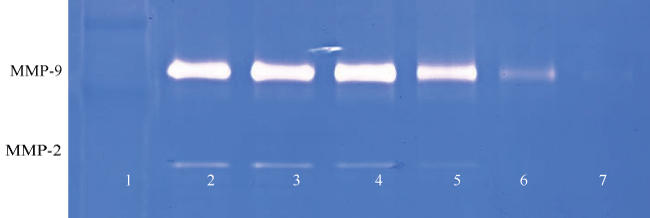
Figure 3E: Gelatinase zymogram of laryngeal squamous cell carcinoma cells treated with a nutrient mixture after PMA stimulation. Legend: 1-Marker, 2-Control/no treatment, 3-7: 10, 50, 100, 500, 1000 μg/ml Nutrient Mixture with 100 ng/ml PMA.
View Figure 3E
.
Figure 3F: Results of densitometry analysis of the FaDu gelatinase zymogram after nutrient mixture treatment shows a dose-dependent decrease in the secretion of MMP-2 and MMP-9.
View Figure 3F
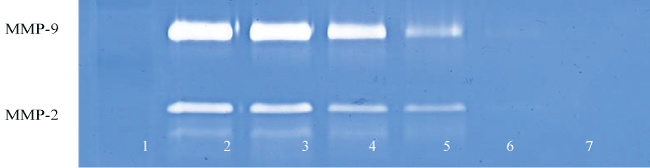
.
Figure 3G: Gelatinase zymogram of tongue carcinoma cells treated with a nutrient mixture after stimulation with PMA. Legend: 1- Marker, 2-Control/no treatment, 3-7: 10, 50, 100, 500, 1000 μg/ml Nutrient Mixture with 100 ng/mL PMA.
View Figure 3G
.
Figure 3H: Results of densitometry analysis of the SCC-25 gelatinase zymogram after nutrient mixture treatment in the presence of PMA showing a dose-dependent decrease in both MMP-2 and MMP-9.
View Figure 3H
![]()
Table 4: Effect of NM with and without PMA on MMP-2 and MMP-9 secretion in laryngeal squamous cell carcinoma (FaDu) and tongue cancer cells (SCC-25).
View Table 4
Discussion
Numerous studies have shown that most cancerous cells secrete massive quantities of matrix metalloproteinases, particularly MMP-2 and MMP-9, which remodel their surrounding microenvironment by dissolving collagen and other extracellular matrix components, thus facilitating metastasis [9,11,22]. Since approximately 90% of cancer fatalities occur as a result of metastasis, the development of effective matrix metalloproteinase inhibitors (MMPI) is a particularly attractive strategy for chemotherapeutic agents. An effective MMPI would not harm healthy cells at clinically relevant concentrations, would have no significant side effects, would target specific cancer cells, and completely suppress the secretion of MMP-2 and MMP-9, thus halting the metastatic process [15,23,24]. Theoretically, the specificity of MMP inhibitors would avoid collateral damage to healthy tissue caused by both chemotherapy and radiotherapy thereby increasing the long-term survival prospects and reducing or eliminating many of the debilitating side effects of current methods of HNSCC therapies [16,17,25].
Experimental and clinical studies have shown a correlation between increased MMPs and tumor progression and metastasis [7]. Moreover, MMP-2 and -9 are identified as useful phenotypic indicators to establish the invasive potential of oral cancers [26-30]. Thus, knowledge of MMP regulation is of importance for developing therapeutic strategies. MMP secretion is regulated at both pre- and post-transcriptional levels. Extracellular factors, including cytokines, growth factors, inducers and inhibitors, have been implicated in the regulation of MMP secretion in different types of tumor cells [31,32]. There are not many studies conducted on HNSCC and TC referring to cytokine and growth factor assays.
In this study, we compared MMP secretion patterns by cytokines (TNF-α, Il-β), inducers (PMA), and mitogenes (LPS) in HNSCC and TC cell lines that express MMP-2 and MMP-9 to different extents. Gelatinase zymography was utilized because of its high sensitivity to gelatinolytic enzymatic activity and ability to detect both pro and active forms of MMP-2 and MMP-9. Among various MMPs, MMP-2 and MMP-9 play a critical role in tumor cell invasion and metastasis [7,33]. MMP-2 (molecular weight 72 kD) is cleaved by other MMPs to yeild the activated forms of 68, 58, and 54 kD and MMP-9 (mol. Wt. 94 kD) into 92 kD. Upon renaturation of the enzyme, the gelatinases digest the gelatin in the gel and reveal clear bands against an intensely stained background. In addition, we also investigated the effect of inhibitors such as doxycycline, dexamethasone, retinoic acid and agents that affect transcription and translation levels such as actinomycin-D and cyclohexamide. The mechanism of doxycycline to inhibit MMP enzymes has been used in various diseases including wound healing, cardiovascular diseases to anticancer properties [34-36]. Similarly, EGCG is shown to inhibit invasion and migration of oral cancer cells by decreased production of MMP-2, MMP-9 and uPA [37,38]. Retinoic acid has been researched as a potential agent for its use in treating different forms of cancer not only in prevention but also in treatment [39,40]. Dexamethasone has been shown to suppress the tumor invasion by targeting NF-κB especially in squamous cell carcinoma [41]. Therefore, we used these agents as comparison. Furthermore, we tested a nutrition mixture that had inhibitory effects on MMP-2 and MMP-9 secretion.
We found that while the FaDu cell line expressed two faint bands the TC cells expressed moderate bands of both MMP-2 and MMP-9. PMA, TNF-α, IL-1β and LPS treatment of both the cell lines strongly increased the secretion of MMP-9 in a dose-dependent manner but had negligible effect on MMP-2. TNF-α had a slight stimulatory effect on MMP-2 while a strong stimulatory effect was observed on MMP-9 in FaDu cells. However, TNF-α had a dose dependent decrease in MMP-9 secretion of TC cells. LPS showed a slight inhibition of MMP-2 and MMP-9 in HNSCC and a slight inhibition of MMP-2 in TC cells. Doxycycline and EGCG inhibited MMP-2 and MMP-9 secretion in a dose-dependent fashion in both the cell lines tested. Similarly, dexamethasone, Actinomycin-D, and cyclohexamide had an inhibitory effect on MMP-2 and MMP-9.
We have developed strategies to inhibit cancer development and its spread using naturally occurring nutrients such as lysine, proline, ascorbic acid, and green tea extract in a nutrient mixture (NM). The NM was designed by focusing on physiological targets in cancer progression and metastasis as documented in clinical and experimental studies. The nutrient mixture was formulated by selecting nutrients that act on critical physiological targets in cancer progression and metastasis as documented in both clinical and experimental studies. Combining these micronutrients expands metabolic targets, maximizing biological impact with lower doses of components. A previous study of the comparative effects of NM, green tea extract (GTE) and EGCG on inhibition of MMP-2 and MMP-9 secretion of different cancer cell lines with varying MMP secretion patterns, revealed the superior potency of NM over GTE and EGCG at equivalent doses [42]. These results can be understood from the more comprehensive treatment offered by the combination of nutrients in NM over individual components of NM since MMP-2 and MMP-9 are mediated by differential pathways.
Our current study demonstrates that the specific mixture of nutrients tested is as effective as the inhibitors doxycycline, dexamethasone, Actinomycin-D, cyclohexamide, and retinoic acid in stopping MMP-2 and MMP-9 secretion in the HNSCC (FaDu) and TC (SCC-25) cell lines. The NM does not harm healthy cells and specifically inhibits overproduction of MMP-2 and MMP-9 in HNSCC. Finally, at a concentration of 500 μg/mL and 1000 μg/mL, the clinically relevant dosages, the NM completely suppressed the secretion MMP-2 and MMP-9 in both cell lines. Furthermore, the NM has been tested in other in vitro studies to prove its safety on vital organs such as, heart, liver and kidney or on the associated serum enzymes [43].
Our results show that inducers, cytokines, and mitogens show downregulation of MMP-2 and MMP-9 in HNSCC and TC. In conclusion, our results showed that cytokines, mitogens, and inhibitors modulated MMP-2 and MMP-9 secretion in FaDu and SCC-25 cells, suggesting the clinical use of MMP inhibitors, especially such potent and non-toxic ones as the nutrient mixture and its component EGCG, in management of head and neck cancers.
Acknowledgement
Dr. Rath Health Foundation, a non-profit organization, funded this research study.
References
-
Howlader N, Noone AM, Krapcho M, Neyman N, Aminou R, et al. (2010) SEER Cancer Statistics Review, National Cancer Institute, Bethesda, MD, based on SEER data submission, posted to the SEER web site 1975-2008.
-
Jemal A, Seigal R, Xu J, Ward E (2016) Cancer statistics, CA Cancer. J Clin 60: 277-300.
-
Maier H, Dietz A, Gewelke U, Heller WD, Weidauer H (1992) Tobacco and alcohol and the risk of head and neck cancer. Clin Investig 70: 320-327.
-
https://seer.cancer.gov/statistics/ctpr.html.
-
Cancer Trends Progress Report.
-
Cooper J, Pajak T, Rubin P, Tupchong L, Brady LW, et al. (1989) Second malignancies in patients who have head and neck cancer incidence effect on survival and implications based on the RTOG experience. Int J Radiat Oncol Biol Phys 17: 449-456.
-
Jones A, Morar P, Phillips D, Field J, Husband D, et al. (1995) Second primary tumors in patients with head and neck squamous cell carcinoma. Cancer 75: 1343-1353.
-
Liotta LA, Tryggvason K, Garebisa A, Hart I, Foltz CM, et al. (1980) Metastatic potential correlates with enzymatic degradation of basement membrane collagen. Nature 284: 67-68.
-
Stetler-Stevenson WG (1990) Type IV collagenases in tumor invasion and metastasis. Cancer Metastasis Rev 9: 289-303.
-
Stetler-Stevenson WG (2001) The role of matrix metalloproteinases in tumor invasion, metastasis and angiogenesis. Surg Oncol Clin N Am 10: 382-392.
-
Fingleton B (2006) Matrix metalloproteinases: roles in cancer and metastasis. Front Biosci 11: 479-491.
-
Charous SJ, Stricklin GP, Nanney LB, Netterville JL, Burkey BB (1997) Expression of matrix metalloproteinases and tissue inhibitor of metalloproteinases in head and neck squamous cell carcinoma. Ann Otol Rhinol Laryngol 106: 271-278.
-
Thomas GT, Lewis MP, Speight PM (1999) Matrix metalloproteinase's and oral cancer. Oral Oncol 35: 227-233.
-
Ruokolainen H, Paaakko P, Turpeenniemi-Hujanen T (2004) Expression of matrix metalloproteinase-9 in head and neck squamous cell carcinoma: a potential marker for prognosis. Clin Cancer Res 10: 3110-3116.
-
Mannello F, Tonti G, Papa S (2005) Matrix metalloproteinase inhibitors as anticancer therapeutics. Curr Cancer Drug Targets 5: 285-298.
-
Matter H, Schudok M (2004) Recent advances in the design of matrix metalloproteinase inhibitors. Curr Opin Drug Discov Devel 7: 513-535.
-
Wallach J, Hornbeck W (2005) Matrix metalloproteinase's and cancer: Novel perspectives in the role and control of matrix metalloproteinases in tumor progression. Biochimie 87: 241.
-
Michi Y, Morita I, Amagasa T, Murota S (2000) Human oral squamous cell carcinoma cell lines promote angiogenesis via expression of vascular endothelial growth factor and upregulation of KDR/flk-1 expression in endothelial cells. Oral Oncol 36: 81-88.
-
Sato T, Sakai T, Noguchi Y, Takta M, Hirakawa S, et al. (2004) Tumor-stromal cell contact promotes invasion of human uterine cervical carcinoma cells by augmenting the expression and activation of stromal matrix metalloproteinase's. Gynecol Oncol 92: 47-56.
-
Pyke C, Kristensen P, Ralfkiaer E, Gröndahl-Hansen J, Eriksen J, et al. (1991) Urokinase-type plasminogen activator is expressed in stromal cells and its receptor in cancer cells at invasive foci in human colon adenocarcinomas. Am J Pathol 138: 1059-1067.
-
Vincenti MP, White LA, Schroen DJ, Benbow U, Brinckerhoff CE (1996) Regulating expression of the gene for matrix metalloproteinase-1 (collagenase): mechanisms that control enzyme activity, transcription and mRNA stability. Crit Rev Eukaryot Gene Expr 6: 391-411.
-
Duffy MJ (1992) The role of proteolytic enzymes in cancer invasion and metastasis. Clin Exp Metastasis 10: 145-155.
-
Chambers AF, Matrisian LM (1997) Changing views on the role of matrix metalloproteinase's in metastasis. J Natl Cancer Inst 89: 1260-1270.
-
Kleainer DE, Stetler-Stevenson WG (1999) Matrix metalloproteinase's and metastasis. Cancer Chemother Pharmacol 43: S42-S51.
-
Fisher JF, Mobashery S (2006) Recent advances in MMP inhibitor design. Cancer Metastasis Rev 25: 115-136.
-
Patel BP, Shah SV, Shukla SN, Shah PM, Patel PS (2007) Clinical significance of MMP-2 and MMP-9 in patients with oral cancer. Head Neck 29: 564-572.
-
Patel BP, Shah PM, Rawal UM, Desai AA, Shah SV, et al. (2005) Activation of MMP-2 and MMP-9 in patients with oral squamous cell carcinoma. J Surg Oncol 90: 81-88.
-
Kato K, Hara A, Kuno T, Kitaori N, Huilan Z, et al. (2005) Matrix metalloproteinase's 2 and 9 in oral squamous cell carcinomas: manifestation and localization of their activity. J Cancer Res Clin Oncol 131: 340-346.
-
Hong SD, Hong SP, Lee JI, Lim CY (2000) Expression of matrix metalloproteinase-2 and -9 in oral squamous cell carcinomas with regard to the metastatic potential. Oral Oncol 36: 207-213.
-
Baker EA, Leaper DJ, Hayter JP, Dickenson AJ (2006) The matrix metalloproteinase system in oral squamous cell carcinoma. Br J Oral Maxillofac Surg 44: 482-486.
-
Egeblad M, Werb Z (2002) New functions for the matrix metalloproteinase's in cancer satoprogression. Nat Rev Cancer 2: 161-174.
-
Folkman J (2002) Role of angiogenesis in tumor growth and metastasis. Semin Oncol 29: 15-18.
-
Kawamata H, Uchida D, Hamano H, Kimura-Yanagawa T, Nakashiro KI, et al. (1998) Active-MMP2 in cancer cell nests of oral cancer patients: correlation with lymph node metastasis. Int J Oncol 13: 699-704.
-
Stechmiller J, Cowan L, Schultz G (2010) The role of doxycycline as a matrix metalloproteinase inhibitor for the treatment of chronic wounds. Biol Res Nurs 11: 336-344.
-
Liu J, Xiong W, Baca-Regen L, Nagase H, Baxter BT (2003) Mechanism of inhibition of matrix metalloproteinase-2 expression by doxycycline in human aortic smooth muscle cells. J Vasc Surg 38: 1376-1383.
-
Hidalgo M, Eckhardt SG (2001) Development of matrix metalloproteinase inhibitors in cancer therapy. J Natl Cancer Inst 93: 178-193.
-
Ho YC, Yang SF, Peng CY, Chou MY, Chang YC (2007) Epigallocatechin-3-gallate inhibits the invasion of human oral cancer cells and decreases the productions of matrix metalloproteinase's and urokinase-plasminogen activator. J Oral Pathol Med 36: 588-593.
-
Koh YW, Choi EC, Kang SU, Hwang HS, Lee MH (2011) Green tea (-)-epigallocatechin-3-gallate inhibits HGF-induced progression in oral cavity cancer through suppression of HGF/c-Met. J Nutr Biochem 22: 1074-83.
-
Mei-Chih Chen, Shih-Lan Hsu, Ho Lin, Tsung-Ying Yang (2014) Retinoic acid and cancer treatment. Biomedicine (Taipei) 4: 22.
-
Jin Man Kim, Sang Wook Kang, Su-Mi Shin, Duck Su Kim, Kyong-Kyu Choi, et al. (2014) Inhibition of matrix metalloproteinase's expression in human dental pulp cells by all-trans retinoic acid. Int J Oral Sci 6: 150-153.
-
Beppu M, Ikebe T, Shirasuna K (2002) The inhibitory effects of immunosuppressive factors, dexamethasone and interleukin-4, on NF-kappaB-mediated protease production by oral cancer. Biochim Biophys Acta 1586: 11-22.
-
SP Netke, MW Roomi, V Ivanov, A Niedzwiecki, M Rath A (2003) Specific combination of ascorbic acid, lysine, proline and epigallocatechin gallate inhibits proliferation and extracellular matrix Invasion of various human cancer cell lines. Emerging Drugs II: IV37- IV50.
-
Roomi MW, Ivanov V, Netke SP, Niedzwiecki A, Rath M (2010) Serum Markers of the Liver, Heart, and Kidney and Lipid Profile and Histopathology in ODS Rats Treated With Nutrient Synergy. Journal of the American College of Nutrition 22: 477.
