

Journal of Sleep Disorders and Management
Marked Increases in Alpha Power Over the Left Prefrontal Region During Days Following Shift Work: A Case Report
Nirosha J. Murugan, Nicolas Rouleau, Lukasz M. Karbowski, Andrew P. Lapointe and Michael A. Persinger*
Behavioural Neuroscience and Biomolecular Sciences Laboratories, Laurentian University, Canada
*Corresponding author:
M. A. Persinger, Biomolecular Sciences, Behavioural Neuroscience and Human Studies Programs, Departments of Biology and Psychology, Laurentian University, Canada, E-mail: mpersinger@laurentian.ca
J Sleep Disord Manag,
JSDM-1-002, (Volume 1, Issue 1),
Case Report
Received: July 30, 2015: Accepted: September 8, 2015: Published: September 10, 2015
Citation: Murugan NJ, Rouleau N, Karbowski LM, Lapointe AP, Persinger MA (2015) Marked Increases in Alpha Power Over the Left Prefrontal Region During Days Following Shift Work: A Case Report. J Sleep Disord Manag 1:002
Copyright: © 2015 Murugan NJ. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Abstract
Quantitative electroencephalographic (QEEG) measurements were completed for a 35 year old of paramedic following two to five days of shift change and rest periods. The most conspicuous and reliable change was a marked increase (factor of 5) in power within the alpha band over the left prefrontal region and, to a lesser degree, increased power within the low-beta band over the right parietal region during the test periods after no work days. These results indicate that regions of cerebral cortices associated with self-monitoring and spatial vigilance are most affected by a schedule that involves serial shifts in sleep schedule but that the structure of employment may attenuate the most significant changes that occur after days of rest. These results are consistent with previous observations that changing sleep and work schedules affect the activity of regions of the human brain that are essential for awareness of spatial content and reasoning.
Keywords
Shift Work, Quantitative EEG, Left Prefrontal Region, Alpha Power, Right Parietal Region
Introduction
Disequilibrium of circadian rhythms secondarily to coerced shifts in work schedules have been known to adversely affect normal sleep characteristics and to increase the probability of diminished vigilance during the days that follow these shifts. Industrial accidents have been shown to occur more frequently during the one or two days after the schedule shift [1]. There is copious literature [2-4] that has shown the effects of schedule shifts on adverse reactions in populations of shift workers.
Considering the types of behaviours that increase the likelihood of an accident after a shift in sleep schedule the most likely region of the human brain to be involved would be the prefrontal lobe, particularly within the left hemisphere. This region is involved with self-monitoring, planning, and inhibitory behaviours [5,6]. The development of modern quantitative electroencephalographic (QEEG) equipment and available software has allowed easy, reliable measurements. Here we present conspicuous disruptions in the stability of cerebral activity within the left prefrontal region for a paramedic who was required to maintain a shifting work schedule.
Methods
The subject who volunteered for this "longitudinal" study was a 35 year old man who reported he had not experienced or sustained any significant medical problems within the previous decade. He had been trained as a paramedic and had begun the traditional procedure of variable shift work as required by the local employer, with no prior exposure to shift-working schedules. Previously, his typical sleep interval was between local midnight and about 06 hr.
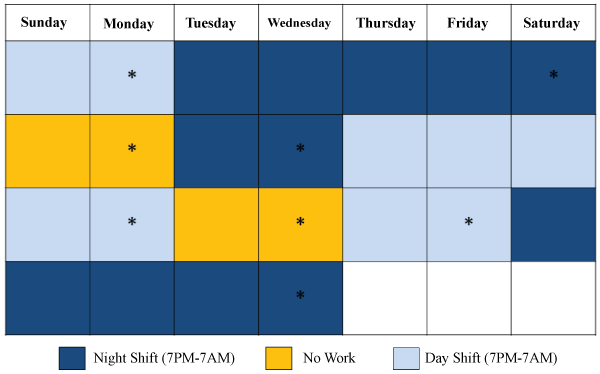
Written consent was obtained from the subject. For each session he was seated in a comfortable chair housed within a darkened, quiet room (an acoustic chamber) and was fitted with a 10-20 EEG cap for about 30 min per session on nine (9) separate occasions over a 25 day period. The days were: Day 1: baseline, Day 2: after 2 days of day shift (7 AM to 7 PM), 3: after 5 days of night shift (7 PM to 7 AM), 4: after 2 days of no work, 5: after 2 days of night shift, 6: after 5 days of day shift, 7: after 2 days of no work, day 8: after 2 days of day shift, and day 9: after 5 days of night shift (Figure 1).
.
Figure 1: The typical work schedule of the paramedic. The asterisks indicate the days where EEG records were obtained. Dark blue indicates days of night shift, light blue indicates days of day shift, and yellow indicates days of no working.
View Figure 1
The procedure was the same for each of the 9 data collection days. QEEG was recorded continuously at 250 Hz (250 samples per second) during the following sequence: 1: 5 min eyes open, 2: 5 min eyes closed, 3: 60 s slow breathing through the nose, 4: 60 s normal breathing, 5: 60 s of a medium rate of breathing through the nose, 6: 60 s of normal breathing, 7: 30 s of hyperventilation through the nose, and, 8: 60 s of normal breathing. At the end of each QEEG episode the short version of the POMS (Profile of Mood States) was administered.
Raw μV data were converted to spectral power (μV/Hz) values within WinEEG. Band ranges including Delta (1.5-4Hz), Theta (4-7.5Hz), Alpha (7.5-14Hz), Beta1 (14-20Hz), Beta2 (20-30Hz), and Gamma (30-40Hz) were selected as default frequency bins within which power spectra were computed. Data were then exported to SPSS v20 and coded by day and the associated preceding shift type for further statistical analyses.
Results
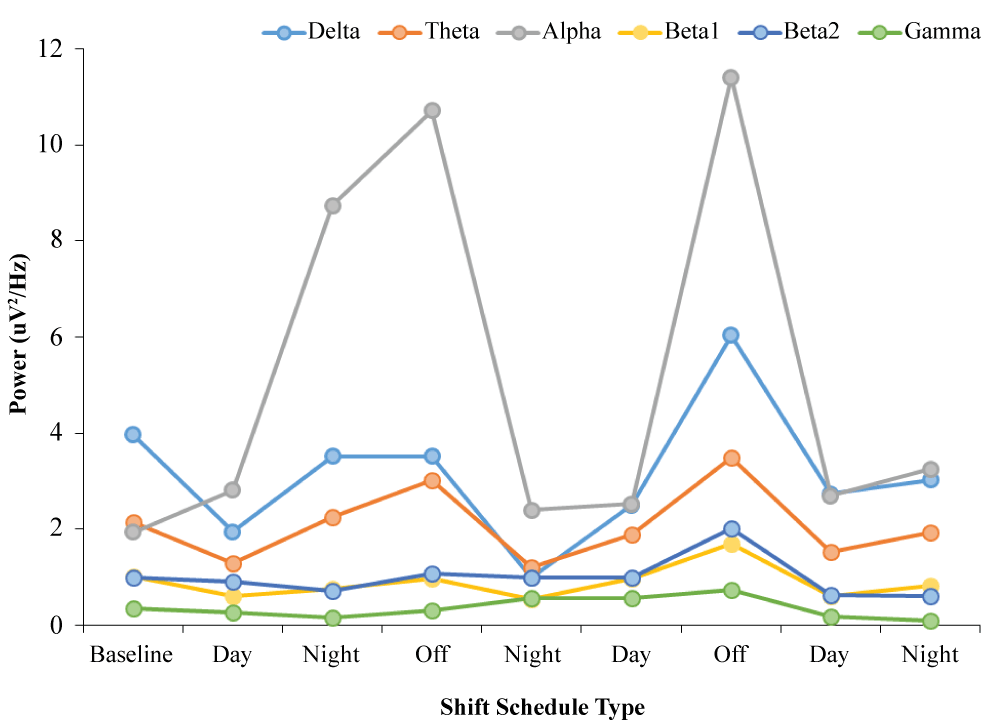
An ANOVA revealed differences in average Alpha (7.5-14 Hz) power over F3 (left prefrontal cortex) during baseline conditions with the participant's eyes open across preceding shift type, F(2,7) = 9.10, p < .05, Ω2 = .78. Post-hoc analyses revealed that the major source of variance was an increase in Alpha power after 2 days of no work (M = 11.07, SEM = .35) relative to values obtained 2 days after day shifts (M = 2.68, SEM = .08), t(3) = -30.18, p < .001). The graphic representation of this effect can be seen in Figure 2.
.
Figures 2: Conspicuous increases in alpha (8-13 Hz) power over the left prefrontal region compared to other frequency bands in a paramedic after 2 to 5 days of shift working times that began during the day or night.
View Figures 2
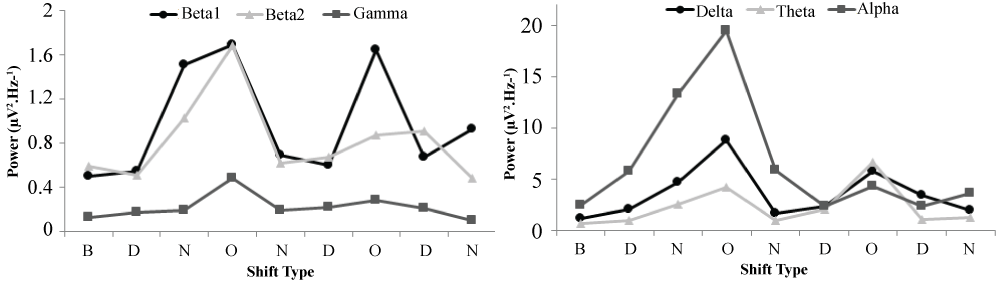
An ANOVA revealed differences in average Beta1 (14-20Hz) power over P4 (right parietal cortex) during baseline conditions with the participant's eyes closed across preceding shift type, F(2,7) = 9.36, p < .05, Ω2 = .79. Post-hoc analyses revealed that the major source of variance was an increase in Beta1 power after 2 days of no work (M= 1.67, SEM =.02) relative to values obtained 2 days after day shifts (M = .61, SEM = .03), t(3) = -23.02, p < .001). After a Bonferroni correction where α = 0.002, these results represent the only statistically significant differences as a function of preceding shift type during the eyes closed baseline condition. These results can be visualized in Figure 3.
.
Figure 3: Conspicuous increases in Beta1 (14-20Hz) power over the P4 channel as a function of time, coded by shift type. Values represented were obtained during baseline (B) measurement, after day shifts (D), after night shifts (N), and after a day off (O).
View Figure 3
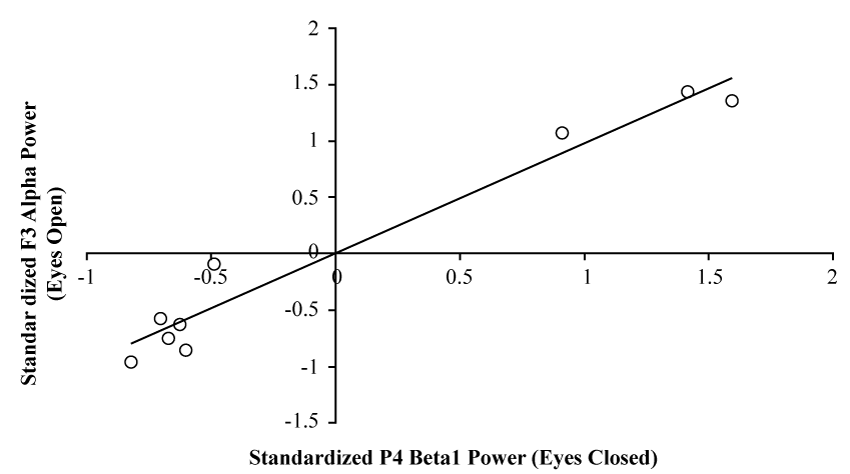
Bivariate, non-parametric correlational analysis of Alpha power over F3 during the eyes open baseline condition (n = 9) and Beta1 power over P4 during the eyes closed baseline condition (n = 9) revealed a strong, positive relationship, r = .98, p < .001, rho = .833, p < .005. These results can be visualized in Figure 4. When examining analogous channels overlying contralateral cerebral structures (i.e. F4 and P3), and correlating spectral power values for the same bands and within the same conditions described above, a similar positive relationship was identified, r = .74, p < .04, rho = .85, p < .005. However, Fisher's z-test revealed a statistically significant difference between Pearson r values resultant of the two correlational analyses (z = 2.24, p < .05), indicating the former strength of the association was greater than the latter.
.
Figures 4: Scatter plot demonstrating the strong relationship between standardized Alpha (7.5-14Hz) power over F3 during the eyes open baseline condition (Y-axis), and standardized Beta1 (14-20Hz) power over P4 during the eyes closed baseline condition (X-axis).
View Figures 4
The specificity of the F3-P4 relationship is emphasized when examining the relationship between global Alpha power during the eyes open baseline condition and global Beta1 power during the eyes closed baseline condition which are demonstrably unrelated, r = .09, p > .05, rho = .37, p > .05. These results can be visualized in Figure 5.
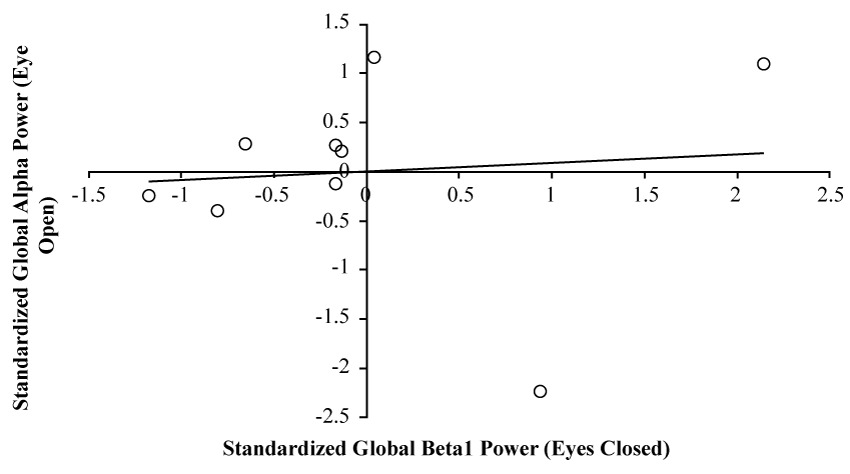
.
Figure 5: Scatter plot demonstrating the non-relationship between standardized, global Alpha (7.5-14Hz) power during the eyes open baseline condition (Y-axis), and standardized, global Beta1 (14-20Hz) power during the eyes closed baseline condition (X-axis).
View Figure 5
Discussion
There has been a general consensus based upon epidemiological data and clinical practice that some people who engage in shift work display alterations in vigilance and increased numbers of behaviours that can contribute to accidents [7,8]. The results of the present case study suggest that the changes in the quantitative electroencephalographic power that occurred over the subject's left prefrontal and right parietal regions may support these behaviours. While the latter is associated with vigilance and relative spatial relationships [9,10], which are important for navigating the social world, the former is associated with self-monitoring of behaviour, organization of thoughts that result in subsequent overt behaviours, and the encoding of experiences into verbal labels [11,12].
That the results are not an artefact of serial measurements and habituation to the measurement setting is indicated by the dominance of the effect following two days of rest. Only the alpha band reflected this component of the shifts in sleeping schedule. The marked increase in alpha power but no changes in the other EEG bands reflects the specificity of influence from altering sleep schedules. Such relative increase in power within the alpha band within the left prefrontal region would be consistent with less proficient activity within this region that could contribute to less accurate self-monitoring and inhibition of distraction particularly within spatial contexts considering the concomitant enhanced activity within the right parietal region.
The markedly reduced magnitude of the right prefrontal and left parietal (the mirror image) strength of association emphasizes the hemispheric lateralization of the effect and reiterates the strong coherence between self-monitoring and input that relates to the spatial-affective environment. It may be relevant that transient increases in alpha power of this magnitude over the left prefrontal region and right parietal region could be expected to increase the subject's proclivity to suggestion [13] and an increased likelihood of increased self-attribution of these experiences.
References
-
Persinger MA, Cooke WJ, Janes JT (1978) No evidence for relationship between biorhythms and industrial accidents. Percept Mot Skills 46: 423-426.
-
Mitler MM, Carskadon MA, Czeisler CA, Dement WC, Dinges DF, et al. (1988) Catastrophes, sleep, and public policy: consensus report. Sleep 11: 100-109.
-
Dinges DF (1995) An overview of sleepiness and accidents. J Sleep Res 4: 4-14.
-
Lyznicki JM, Doege TC, Davis RM, Williams MA (1998) Sleepiness, driving, and motor vehicle crashes. Council on Scientific Affairs, American Medical Association. JAMA 279: 1908-1913.
-
Siddiqui SV, Chatterjee U, Kumar D, Siddiqui A, Goyal N (2008) Neuropsychology of prefrontal cortex. Indian J Psychiatry 50: 202-208.
-
Goldman-Rakic P.S (1996) The prefrontal landscape: implications of functional architecture for understanding human mentation and the central executive. Philos Trans R Soc Lond B Biol Sci 351: 1445-1453
-
Horne J, Reyner L (1999) Vehicle accidents related to sleep: a review. Occup Environ Med 56: 289-294.
-
Scott LD, Rogers AE, Hwang WT, Zhang Y (2006) Effects of critical care nurses' work hours on vigilance and patients' safety. Am J Crit Care 15: 30-37.
-
Corbetta M, Shulman GL, Miezin FM, Petersen SE (1995) Superior parietal cortex activation during spatial attention shifts and visual feature conjunction. Science 270: 802-805.
-
Hilgetag CC, Théoret H, Pascual-Leone A (2001) Enhanced visual spatial attention ipsilateral to rTMS-induced'virtual lesions' of human parietal cortex. Nat Neurosci 4: 953-957.
-
Gabrieli JD, Poldrack RA, Desmond JE (1998) The role of left prefrontal cortex in language and memory. Proc Natl Acad Sci U S A 95: 906-913.
-
Stuss DT, Benson DF (1984) Neuropsychological studies of the frontal lobes. Psychol Bull 95: 3-28.
-
Williams JD, Gruzelier JH (2001) Differentiation of hypnosis and relaxation by analysis of narrow band theta and alpha frequencies. Int J Clin Exp Hypn 49: 185-206.
